Neurophysiology
Assoc. Prof Craig Johnson, IVABS, Massey University
Key points
■ Myelination increases the speed at which impulses are propagated along the myelinated fibre by enabling saltatory conduction.
■ Action potentials trigger the release of neurotransmitter at a chemical synapse or neuromuscular junction. The neurotransmitter crosses the synaptic cleft and binds to receptors on the postsynaptic membrane causing chemically-gated ion channels to open. Ion transfer causes change in the postsynaptic membrane potential.
■ Neurotransmitters are classed as excitatory or inhibitory depending on whether they depolarise or hyperpolarise the postsynaptic potential.
There are many different types of neurons, all of which are designed to perform different functions, yet all types work using the same principles and components. Information processing occurs within the components of the neuron (Fig. 3.5) as well as at the connections between neurons. The functional capacity of the central nervous system to process information is due to both the complexity of individual neurons and their connections. Most types of neuron only connect with other neurons. Some types connect with sensory receptors, while others connect with effector tissues such as muscle of the body, viscera, blood vessels and glands.
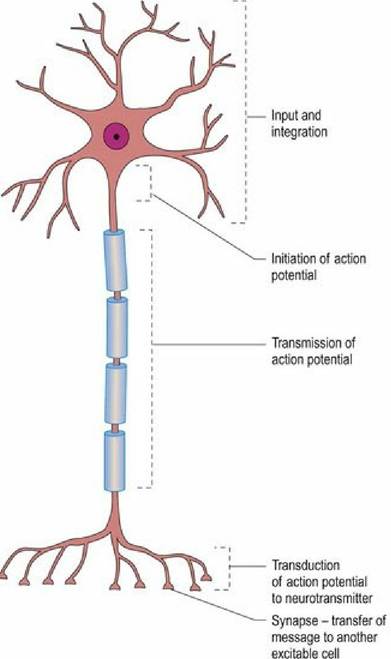
Fig. 3.5 Functional regions of a neuron.
Membrane potential of excitable cells
The cell membrane is composed of a lipid bilayer that has a hydrophilic surface and a hydrophobic interior. Embedded in the cell membrane are proteins such as molecular pumps and ion channels; these are important for electrical activity. The cell membrane is responsible for the generation and maintenance of the membrane potential. In addition, the membrane of excitable cells can propagate an action potential.
The membrane potential is formed by concentration gradients of ions across the plasma membrane. The gradient is formed by the high resistance of the lipid bilayer and the action of ion channels embedded in the cell membrane (Fig. 3.6).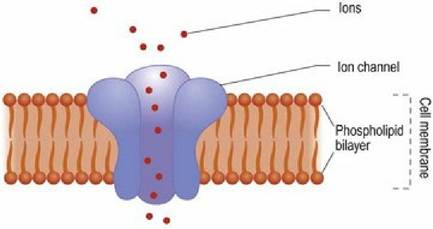
Fig. 3.6 The ion channel spans the lipid bilayer and, when open, permits ions to flow between the inside and the outside of the cell.
The generator of the membrane potential is the molecular co-transporter sodium potassium ATPase pump (Fig. 3.7). This molecule uses chemical energy in the form of ATP to transport sodium ions out of cells and potassium ions into cells. For every three positively charged sodium ions removed from the cell, two positively charged potassium ions replace them. These ionic gradients generate an electrical potential difference across the membrane, called membrane potential. The size of a cell’s membrane potential depends on how permeable that cell’s membrane is to particular ions. Permeability is a consequence of the number and type of ion channels. Ion channels allow ions to diffuse across the plasma membrane, down their concentration gradient. For example, a smooth muscle cell typically has a membrane potential of -50 mV whereas that of striated muscle will be about -95 mV. The membrane potential for a particular cell can be calculated using the Goldman equation. This equation takes into account the concentrations of ions inside and outside the cell, and the permeability of that cell membrane to those ions.
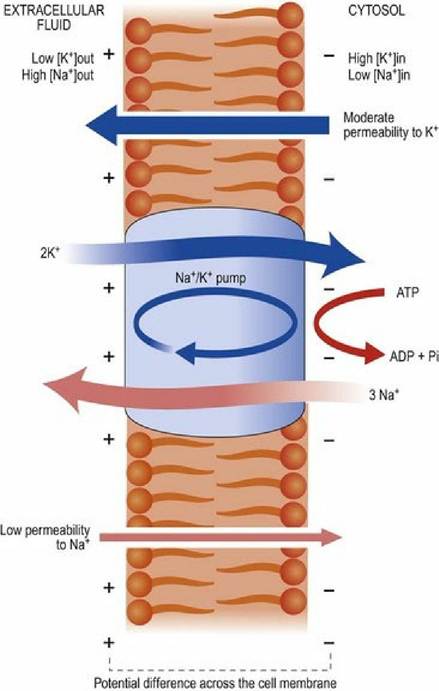
Fig. 3.7 Membranes have a default permeability to ions. The sodium-potassium ATPase pump establishes ion concentration gradients across the plasma membrane; this is the basis of the resting membrane potential.
Cell membranes have an inherent permeability to ions such as sodium and potassium. The ATPase pump establishes a concentration gradient of ions, such that the ions diffuse down their concentration gradient until the charge thus generated balances the osmotic force of the concentration gradient.
Action potential
An action potential is generated when the membrane potential of the axon hillock (see Fig.
3.2) depolarises sufficiently to reach threshold potential. When threshold potential is reached, voltage-gated sodium and potassium channels open allowing massive ion diffusion across the membrane. Sodium ions move more rapidly than potassium ions and so for a brief period the membrane potential becomes much less negative as positively charged sodium ions flow into the cell. The further reduction in membrane potential allows even more voltage-gated channels to open. The change in membrane potential spreads to adjacent areas of the plasma membrane and they, too, pass threshold and become highly permeable to sodium and potassium ions. Eventually the neuron reaches a point where the membrane potential is reversed and the voltage-gated channels begin to close. The closure of the voltage-gated ion channels brings the movement of ions to an end and the membrane potential equilibrates to normal values. This change in membrane potential continues to spread along the length of the axon and is the basis for the electrical conduction of information along nerves (Fig. 3.8).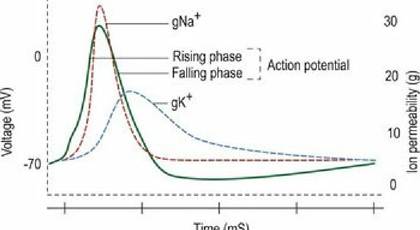
Fig. 3.8 Schematic diagram illustrating the generation of an action potential owing to the changing
permeability of the cell membrane to Na+ and K+.
Myelination and nerve conduction
The rate at which an action potential travels along an axon is influenced by many factors including the diameter of the axon and the number of ion channels in the plasma membrane. Many axons are wrapped in a layer of lipid-rich myelin. Myelin prevents ion exchange in the internodal areas and consequently these sections of axon have few ion channels and molecular pumps. Thus action potentials are generated only at the nodes of Ranvier. In myelinated axons, action potentials travel by jumping from one node of Ranvier to the next in a process called saltatory conduction. This makes action potential conduction both much faster (nerve impulses are conducted at speeds of up to 120 m/s) and more energy efficient.
In non-myelinated axons, ion channels and molecular pumps are spread along the length of the axon. This reduces the speed of action potential conduction as well as increasing the energy expenditure.Synapses
Neurons can be connected to each other in two distinct ways:
1. Chemical synapses in which a chemical neurotransmitter bridges the 30-50 nm space between the pre- and postsynaptic membrane (Fig. 3.9).
2. Electrical synapses in which gap junctions (protein pores) form physical contact between cells. They allow faster conduction of membrane potential changes between neurons. Unlike chemical synapses they cannot amplify signals and are usually bidirectional. They are common in invertebrates and are found in the retina and cerebral cortices of vertebrates.
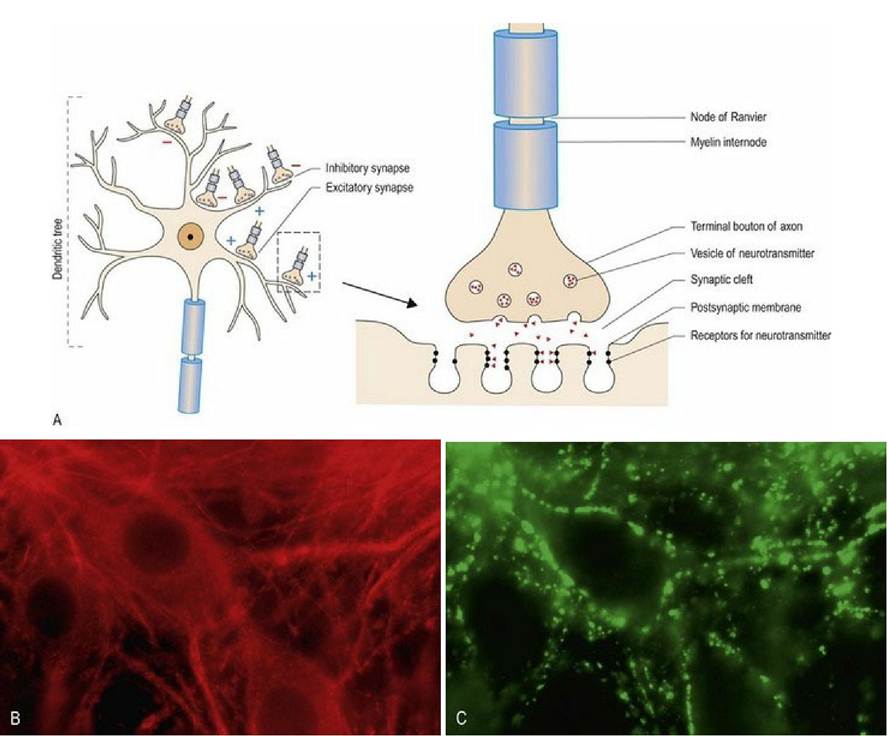
Fig. 3.9 Chemical synapse. (A) The electrical energy of the action potential is transmitted between adjacent neurons, or from neurons to muscle cells, by the release of chemical neurotransmitters. (B,C) Cultured murine spinal cord neurons colabelled for tubulin (red) a neuronal cytoskeletal protein and synaptophysin (green) to identify synapses. The neuronal nucleus can be seen as a negatively stained shadow in the centre of the neuronal cell bodies in figure 9B; it is approximately 7-8 μm in diameter (reproduced with permission from Thomson et al, 2008).
The vast majority of the synapses in the mammalian brain are chemical synapses; the neuromuscular junction is also a chemical synapse.
When an action potential arrives at a chemical synapse, it stimulates release of a chemical neurotransmitter from the axonal terminus into the synaptic cleft. This neurotransmitter binds to receptors on the postsynaptic membrane, stimulating chemically gated ion channels. These channels open and induce ion fluxes changing the postsynaptic membrane potential. Depending on its location in the nervous system, the recipient neuron can receive tens or even hundreds of thousands of inputs from other neurons; each one can induce changes in the local membrane potential.
The numerous changes are collated and the recipient cell processes all the incoming information. The collated input may stimulate the recipient cell to depolarise or not. If it depolarises, then information continues to be spread through the nervous system.Under normal circumstances, information processing does not occur at the neuromuscular junction. Thus the arrival of an action potential at the axonal terminus activates calcium channels resulting in the release of neurotransmitter into the synaptic cleft and an action potential in the postsynaptic muscle fibre and muscle contraction. The number of action potentials arriving at the axonal terminus determines the strength of the muscle contraction.
Excitatory and inhibitory neurotransmitters
Neurotransmitters are chemicals that transmit nerve impulses across a synapse. They can be either inhibitory or excitatory. Excitatory neurotransmitters open ion channels allowing extracellular positive ions, such as sodium, to diffuse into the cell. This makes the inside of the cell less negative, thereby decreasing the potential difference across the membrane. An example of an excitatory neurotransmitter is glutamate. Inhibitory neurotransmitters open ion channels permitting extracellular negative ions, such as chloride, to diffuse into the cell, or intracellular positive ions such as potassium to diffuse out. The inside of the cell becomes more negative and increases the potential difference existing across the membrane. In the brain the major inhibitory neurotransmitter is GABA, while in the spinal cord it is glycine. An individual neuron will generally release only one type of neurotransmitter molecule, thus neurons can be termed excitatory or inhibitory.
The generalised neuron
Even though there is a huge variety in the shape and degree of connectivity of different neurons, the concept of the generalised neuron covers most of the variations and is a useful way to describe the concepts of nerve function.
Neurons process incoming information and may generate and propagate electrical charges as follows. The neuron receives information from many other neurons via chemical or electrical synapses. These inputs cause local fluctuations in membrane potential at the postsynaptic membrane. The changes in the membrane potential spread over the surface of the neuron. The dendritic processes of the neuron together with the cell body integrate all of the changes in membrane potential from all regions of the neuron’s sensory field. The summated changes affect the membrane potential at the axonal hillock. This is the critical area for the propagation of information. If this area reaches the threshold potential for the neuron, then an action potential will develop, spread along the axon to its synaptic termination where it will stimulate, or inhibit, the next neuron in the chain.Information processing within and between neurons
The primary function of the nervous system is to receive inputs, process information and produce the appropriate output. The processing of information is carried out at several levels. Individual neurons process information from their receptive field to produce changes in the rate at which they generate action potentials. Neurons are interconnected into groups that work together to carry out specific processing tasks. Groups of neurons are interconnected into systems that deal with specific functions and systems interconnect at progressively higher levels to produce an animal’s nervous system. This multi-level, modular design allows powerful and rapid information processing that enables the organism to sense its environment and behave in a manner appropriate to its perception. This modular concept is explained further in Chapter 4.