Supporting structures
Assoc. Prof Craig Johnson, IVABS, Massey University
Key points
■ The brain and spinal cord are surrounded by three layers of meninges - the dura mater, arachnoid mater and pia mater.
■ Cerebrospinal fluid is produced by choroid plexi in the ventricular system of the brain. Its flow is largely rostral to caudal in the ventricular system and subarachnoid space from which it is drained.
■ The CNS receives 20% of the total cardiac output, with the metabolically active grey matter receiving more than the white matter.
■ The arterial blood supply to the brain is based around four pairs of arteries arising from the cerebral arterial circle, and one pair originating from the basilar artery. There are major, clinically important, species differences in the pattern of this arterial supply.
■ The blood-brain barrier surrounding blood vessels is selectively permeable to bloodborne components. This protects the CNS from microorganisms but also limits drug permeability. However, CNS disease can compromise the function of blood-brain barrier.
Meninges
The brain and spinal cord are surrounded by three layers of connective tissue called meninges (Fig. 3.10). From superficial to deep, these are the dura mater, the arachnoid mater and the pia mater (meninx - Gk = membrane; dura - L = tough; mater - L = mother; arachnoid - Gk = cobweb like; pia - L = tender).
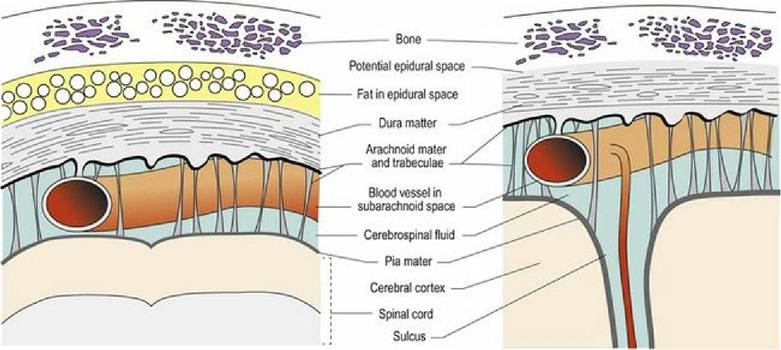
Fig. 3.10 Sections through meninges surrounding the spinal cord (left) and brain (right).
The dura mater is composed of a thick layer of fibrous tissue, lined by epithelium and is also known as the pachymeninx (pachys - Gk = thick). The arachnoid mater is a thin membrane that encloses the cerebrospinal fluid (CSF) in the subarachnoid space. The arachnoid mater has numerous, fine filaments that traverse the subarachnoid space connecting with the pia mater (see Figs.
3.11B and 7.1). The pia mater is fused to the surface of the neuraxis. The arachnoid and pia mater comprise the leptomeninges (lepto - Gk = thin). The epidural space exists in the spinal canal and contains epidural fat and blood vessels. Inside the neurocranium, the dura is fused to the periosteum and the epidural space is a potential space only. The dura and arachnoid mater are fused in both regions; the subdural space is also a potential space. Potential spaces may be opened forming an actual space by pathological processes such as haemorrhage.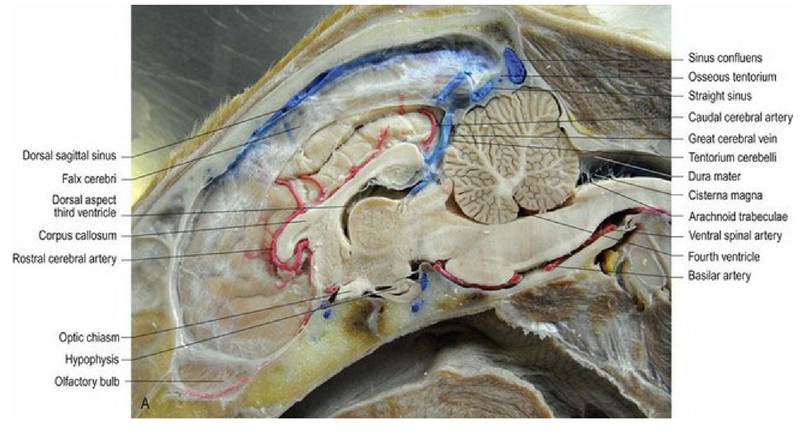

Fig. 3.11 (A) Dog head, perfusion fixed and with latex injected into the vasculature. This is a paramedian section, cut just to the left of the midline, to include the falx cerebri, thus the ventral aspect of the third ventricle and the mesencephalic aqueduct are not visible. (B) The subarachnoid space extends rostrally from the transverse fissure into the caudal diencephalic region. (Specimens courtesy of Mr. Allan Nutman, IVABS, Massey University.)
In two places, the dura mater folds inwardly to form double-layered curtains that hang from the roof of the cranial vault. The falx cerebri runs longitudinally separating the two cerebral hemispheres. The tentorium cerebelli runs transversely separating the caudal poles of the hemispheres from the rostral aspect of the cerebellum (Fig. 3.11A).
The meninges extend around the spinal nerve roots as sleeves and may reach as far peripherally as the intervertebral foramina (Fig 1.1). At this level, the meninges fuse with the epineurium to form distinct cuff zones. At the cuff zones, drugs can diffuse more easily into the CNS. This characteristic is utilised in epidural anaesthesia to block nerve conduction in the spinal roots.
The ventricular system
The ventricular system is derived from the fluid-filled centre of the embryonic neural tube.
In the brain, it comprises four expanded regions forming ventricles, interconnecting foramens and tubes (see Figs. A4, A5, A9-27). There is a curved lateral ventricle oriented longitudinally in each cerebral hemisphere that is connected to the third ventricle via an interventricular foramen. The third ventricle is a dorsoventrally oriented ring, surrounding the interthalamic adhesion. It connects via the mesencephalic aqueduct to the fourth ventricle located ventral to the cerebellum. Caudally, the dorsal aspect of this ventricle closes over at the obex to form the spinal canal. Laterally, the fourth ventricle is connected via two lateral apertures to the subarachnoid space. The ventricular system and the subarachnoid space contain cerebrospinal fluid (CSF) (Fig. 3.12).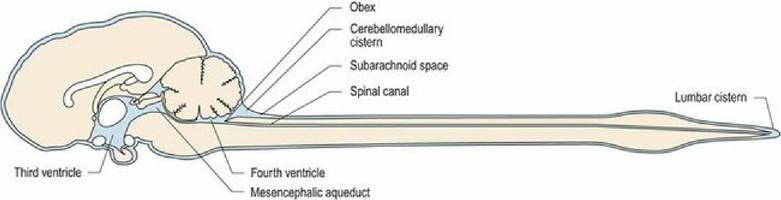
Fig. 3.12 Median section of the canine brain, ventricular system and subarachnoid space of the CNS. The
lateral ventricles are lateral to the plane of this image.
Cerebrospinal fluid: production and circulation
Cerebrospinal fluid (CSF) is produced by small arteries and arterioles in the leptomeninges and the choroid plexuses. A choroid plexus is a collection of capillaries interposed in the choroid membrane (tela choroidea). The tela choroidea forms where the pia mater apposes the ependyma with no intervening neural tissue. There is a choroid plexus in each of the four ventricles of the brain. Cerebrospinal fluid is produced by ultrafiltration of blood plasma through the choroid plexus and modified by secretions from the epithelial secretory cells. The CSF provides physical and chemical protection for the CNS. The endothelial-epithelial barrier between the blood and CSF is only selectively permeable to bloodborne substances.
The CSF flows from the lateral ventricles, through the interventricular foramens to the third ventricle, caudally through the mesencephalic aqueduct to the fourth ventricle.
From here some flows down the central canal of the spinal cord. The majority of CSF leaves the central space through lateral apertures of the fourth ventricle and enters the sub-arachnoid space. The CSF is drained from the sub-arachnoid via three main routes: (a) by the venules of the sub-arachnoid space; (b) into the venous sinuses of the brain through invaginations in the wall of the sinus called arachnoid villi; and (c) by the lymphatic vessels of the cranial and spinal nerve roots.Blood supply
Although the CNS accounts for only 2% of the body weight, it has a disproportionately high metabolic rate and receives about 20% of the total cardiac output. The oxygen requirements of the synapses and neuronal cell bodies are greater than those of the axons, thus the grey matter receives more blood flow than the white matter. Additionally, association/integration areas have greater requirements than other areas and so the forebrain is more vascular than other CNS regions.
Arterial blood supply of the brain is based around five pairs of arteries (Figs. 3.13-15, A3). Four of these arise from the cerebral arterial circle located on the ventral surface of the forebrain. These arteries include the rostral, middle and caudal cerebral arteries and the rostral cerebellar artery. The caudal cerebellar arteries originate from the basilar artery. The basilar artery is continuous with the ventral spinal artery at the foramen magnum. It runs longitudinally on the ventral aspect of the brainstem and connects with the caudal aspect of the arterial circle. The major brain arteries traverse the surface of the brain in the sulci, and send smaller branches to perfuse the deep tissues. Whilst there are anastomoses between the arteries on the surface of the CNS, the penetrating arteries do not communicate with each other and so blockages of individual end-arteries lead to ischaemia of defined parts of the CNS.
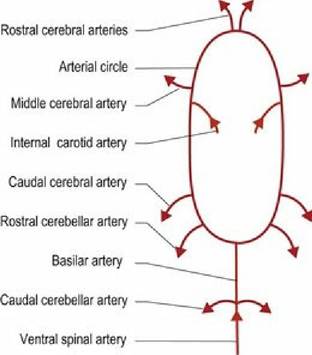
Fig.
3.13 Schematic of the blood supply to the brain of the dog.
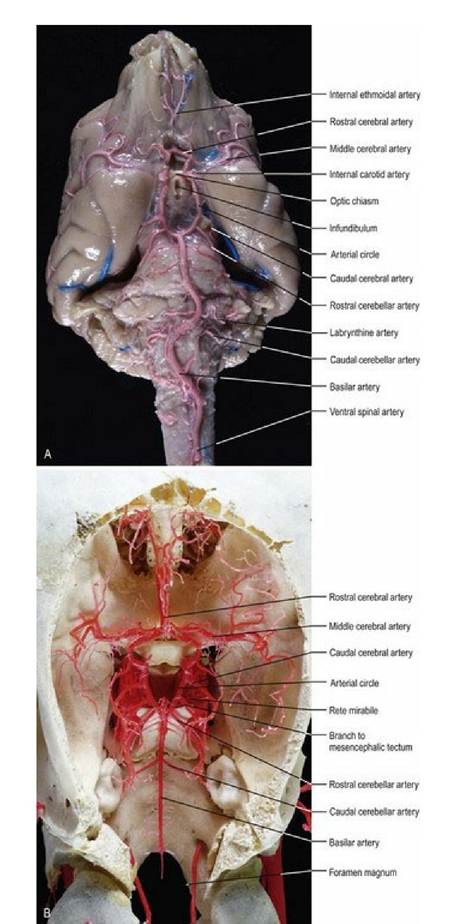
Fig. 3.14 The arterial supply of the dog and sheep brain. The vessels of the dog brain have been infused with latex, while the sheep vasculature is a resin cast. (Specimens prepared by Mr. Allan Nutman, IVABS, Massey University.)
_ Maxillaryartery Retemirabile Basilarartery
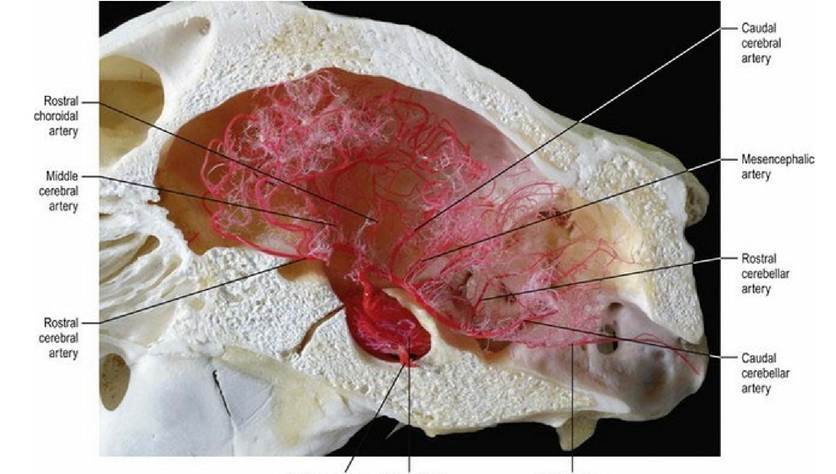
Fig. 3.15 The rete mirabile of the sheep brain. (Vascular cast prepared by Mr. Allan Nutman, IVABS,
Massey University.)
Although there are some species differences in the arteries of the brain, these are relatively minor. However, there are major differences in how blood gets to the brain; these differences are clinically important. Generally speaking there are four possible routes by which the blood may arrive at the arterial circle; the internal carotid artery, the basilar artery, the maxillary artery and the vertebral artery. The common domestic animals have various combinations of these routes. The maxillary artery and the vertebral artery may have rete mirabile (rete - L = net, mirabile - L = marvellous) where the major vessel breaks into many convoluted small branches before re-anastomosing. The rete mirabile may act as a heat exchanger to protect the brain from major temperature changes and act to reduce pulsations in the blood perfusing the CNS. The cat, sheep (Fig. 3.15) and ox have one, or more, rete mirabile.
Patterns of arterial supply to the brain
1. General form (dog, human). The internal carotid artery and the basilar artery both carry blood to the cerebral arterial circle. Carotid blood reaches most of the cerebral hemispheres except the caudal portion. Vertebral blood supplies the rest of the brain (Fig. 3.16).
2. Sheep and cat. The proximal two-thirds of the internal carotid artery is absent in the adult and the direction of blood flow is caudad in the basilar artery.
The only supply of blood to the cerebral arterial circle originates from the maxillary arteries via a rete mirabile. Maxillary blood supplies all the brain except the caudal part of the medulla.3. Ox. As in the sheep and cat the proximal two-thirds of the internal carotid is absent in the adult and the direction of blood flow is caudad in the basilar artery. Unlike the sheep there are two anastomosing branches, one from the maxillary artery and one from the vertebral artery, both of which have rete mirabile. Maxillary and vertebral blood supplies the whole of the brain.

Fig. 3.16 The blood supply to brains of different species varies; (A) dog and humans, (B) sheep and cat, (C) ox. Magenta represents supply from the internal carotid artery; yellow from the vertebral artery; blue from the maxillary artery and green is from both the vertebral and maxillary arteries.
Blood supply and animal slaughter
Animal welfare legislation in many countries prohibits slaughter without prior stunning in order to ensure that animals do not perceive pain during throat cutting and exsanguination. Cutting the throat cuts the common carotid artery and jugular veins. The teaching of some religions does not allow for stunning to take place prior to slaughter and legislation in some countries provides them with an exemption from the general requirement for stunning. In these animals that are not stunned there will be a period of time after the throat cut and before the onset of insensibility. It has recently been shown that the animal may perceive pain during this period. The duration of sensibility following the throat cut will depend on whether the vertebral arteries can keep the brain supplied with oxygen. This period varies between species because of anatomical differences. It is thought that the period of sensibility is between 8 and 20 seconds in the sheep, but between 34 and 85 seconds in cattle.
Arterial supply to the spinal cord
The spinal cord is supplied segmentally by spinal branches arising from the vertebral artery and aorta, and entering the spinal canal at the intervertebral foramens. Within the spinal canal, each branch divides into a smaller dorsal, and larger ventral branch. The dorsal branches enter the spinal cord with the dorsal root. The ventral branches unite to form the unpaired, median, ventral spinal artery.
Venous drainage of the CNS
The neuraxis and meninges are drained by veins and sinuses (see other texts for details of the veins). In sinuses, valves are absent or poorly developed, hence retrograde blood flow can occur. Sinuses in the brain form a dorsal and ventral set. The dorsal set is primarily midline with the dorsal sagittal sinus, sited in the falx cerebri draining caudally to meet the straight sinus at the confluences of the sinuses located at caudal end of the falx cerebri. The straight sinus drains the great cerebral vein from the deeper cerebrum. Drainage from the confluence is laterally into the ventrally directed, paired transverse sinuses, beginning in the dorsal part of the occipital bone. The transverse sinus terminates by dividing into the sigmoid and temporal sinuses in the ventrocaudal aspect of the skull. The ventral set of sinuses begins with the paired cavernous sinus lying subdurally on the floor of the rostral fossa, surrounding the hypophysis. The cavernous sinus receives venous drainage from the ophthalmic plexus, and may, depending on the species, interconnect across the midline by the intercavernous sinus. The cavernous sinuses drain caudally into the ventral petrosal sinus that connects to the sigmoid sinus. The dorsal petrosal sinus drains the basal vein of the cerebrum into the transverse sinus. The sinuses drain into the maxillary, vertebral, occipital and internal jugular veins (Fig. 3.17).
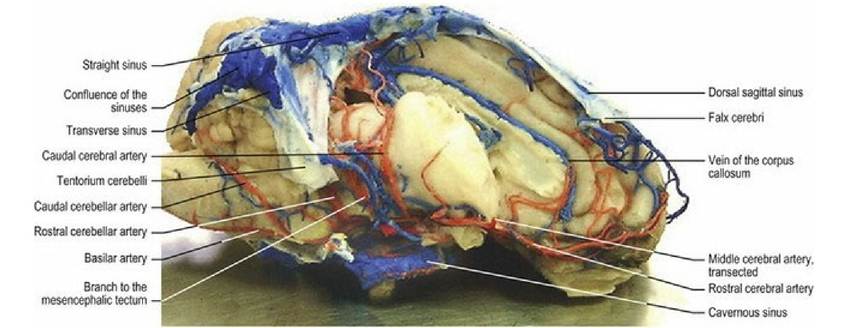
Fig. 3.17 Sheep brain lateral aspect, vasculature filled with latex (red for arteries and blue for veins and
sinuses). The right hemisphere has been removed to show deeper vessels. (Specimen prepared by Mr. Allan Nutman, IVABS, Massey University.)
The spinal cord has an internal vertebral venous plexus lying on the floor of the spinal canal, with paired vessels that diverge to drain into intervertebral veins at each intervertebral foramen (Fig 3.18).
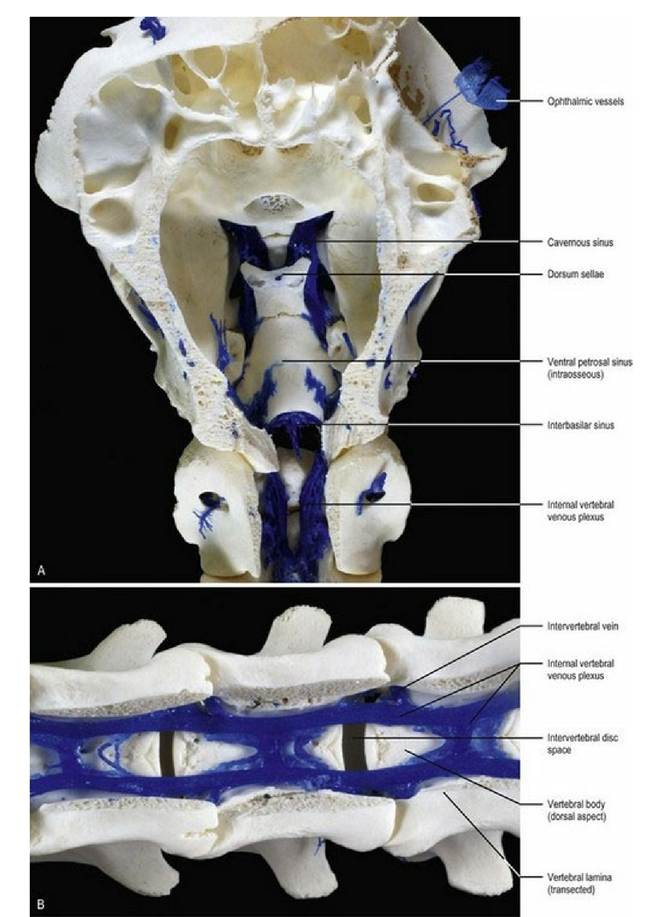
Fig. 3.18 Resin casts of the venous sinuses and veins on the ventral aspect of the neuraxis in the sheep. (A) Base of the neurocranium of sheep, dorsal aspect. (B) Base of the spinal canal of sheep, dorsal aspect. (Specimens prepared by Mr. Allan Nutman, IVABS, Massey University.)
Blood-brain barrier
The blood-brain barrier (together with the blood-CSF barrier) prevents some molecules that may be present in the blood from diffusing into the CNS parenchyma. This protective mechanism aims to limit the exposure of the CNS tissues to pathogens and molecules that could have a deleterious effect. Because of these barriers, the composition of the CSF and the extracellular fluid in the CNS is different to that in the rest of the body.
Structurally the blood-brain barrier has three components: (a) tight junctions between the capillary endothelia; (b) a thick basement membrane; and (c) a layer of astrocyte foot processes that surround the capillaries (Fig. 3.19). The barrier is highly permeable to water, carbon dioxide, oxygen and lipid-soluble substances, and less permeable to electrolytes. Carrier molecules mediate transport of glucose and some other small molecules. It is almost totally impermeable to proteins and larger molecules, especially those that bear an electrical charge. Some areas, such as the hypothalamus, the area postrema and pineal gland, lie outside of the blood-brain barrier. These areas contain sensors for osmolarity, glucose concentration and pH, and send information into regulatory areas of the CNS. The blood-brain and blood-CSF barriers function to maintain a highly controlled CNS environment, but also have clinical significance with respect to medical treatment of the CNS as they will exclude many drugs. However, the blood-brain barrier in developing and senescent animals is more porous than that of mature adults.
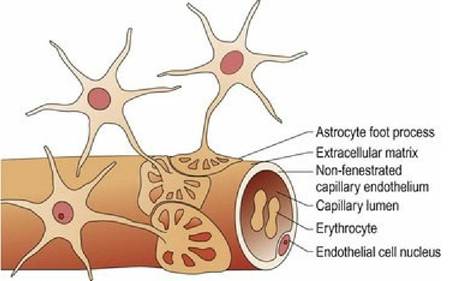
Fig. 3.19 The blood-brain barrier.
Examples of drugs that cannot normally cross the blood-brain barrier include some antibiotics and neuromuscular blocking agents. However, pathological processes, such as inflammation, can compromise the function of these barriers allowing entry of compounds that are normally excluded.