Nociception
Key points
■ A noxious stimulus may be sensed by specific nociceptors, or non-specifically by other receptor types.
■ A noxious stimulus may stimulate reflex activity, or it may be transmitted to the brain by pathways in all funiculi and on both sides of the cord.
■ Pain is the conscious perception of a noxious stimulus; it is accompanied by an unpleasant emotional response.
■ Loss of conscious perception of noxious stimuli in the body caudal to a spinal cord lesion implies severe destruction of the spinal cord.
■ Noxious stimuli in the head are conveyed by CN V to the pontine nucleus of CN V for reflex function. Fibres also travel rostrally to the contralateral somatosensory cortex for conscious perception.
■ Noxious stimuli from the viscera are transmitted to the CNS primarily via sympathetic and parasympathetic fibres. They may induce local reflex activity, autonomically mediated changes and conscious perception.
Nociception (nocere - L = to injure), is defined as the sensory modality that is preferentially triggered by a noxious stimulus. The stimulus can be chemical, thermal or mechanical. It may trigger a variety of somatic or autonomic responses and reflexes, and may also result in the experience ofpain.
Pain is defined by the International Association for the Study of Pain (http://www.iasp-pain.org) as ‘An unpleasant sensory and emotional experience associated with actual or potential tissue damage...’.
Noxious stimuli generate action potentials in a variety of sensory pathways, but only when those stimuli are received and integrated in the cerebrum, and associated with an unpleasant emotional status, may they be considered to be causing pain. Thus pain perception has both a sensory and emotional component.
‘The inability to communicate verbally does not negate the possibility that an individual is experiencing pain’ (http://www.iasp-pain.org).
The non-verbal animal’s way of conveying pain is through pain-related behaviour such as avoiding the noxious stimulus, depression/withdrawal or self-selection of analgesia as observed in laboratory animals.Noxious stimuli may stimulate a variety of receptors, some of which respond specifically to noxious (tissue-damaging) stimuli. Other receptors, such as tactile mechanoreceptors, primarily respond to mechanical distortion by non-noxious stimuli, but will also respond to the higher-intensity, noxious stimuli.
Nociceptive input from body
Approximately 50% of afferent axons in cutaneous nerves are associated with nociception. Noxious stimuli are transmitted via two types of fibres in the PNS. Aδ-fibres transmit pinprick pain. As they are myelinated, they are rapidly conducting and transmit ‘fast pain’. Transmitted stimuli are localisable to a body area, warn the brain of the potential for tissue damage and cause rapid responses by the brain, for example, promoting activities that remove the body from the noxious stimulus. C-fibres transmit acute pain, but much more slowly as they are non-myelinated. In humans, C-fibre stimulation causes the aversive aspects of pain described as dull, aching, throbbing, burning, depression or nausea. Such pain causes the individual to withdraw and rest aiming to promote healing. A clinical example of Aδ-fibre transmitted stimulus is light squeezing of the skin using haemostats whereas C-fibres are stimulated by the crushing pain that can be produced by the squeezing of the digits with a strong pair of haemostats. Aδ-fibre stimuli are readily localised, whereas C-fibre stimuli may be poorly localised by the brain.
Both fibre types enter via the dorsal root and synapse in the dorsal horn. From there, connections are made within or between segments, for reflex activity such as the withdrawal reflex, in which the animal withdraws the limb from the noxious stimulus. Simultaneously, in the standing animal, the contralateral limb will be stimulated reflexively to extend, but in the normal, recumbent animal, the crossed extension reflex is suppressed.
The noxious stimulus will also be transmitted cranially in a variety of pathways found in all funiculi. For conscious perception, the tracts ultimately project via the thalamus to the somatosensory cortex. Localisation of pinprick (fast) pain to the body surface may involve the post-synaptic tract in the dorsal funiculus. This tract conveys both noxious and non-noxious stimuli from the skin and arises from incoming sensory nerves that have synapsed in the dorsal horn. Fibres run cranially mingled with those conveying discriminative touch in the fasciculus gracilis and cuneatus. Other types of noxious stimuli causing stimulation of Aδ- or C-fibres may use tracts in the lateral or ventral funiculi such as the spinothalamic tract, spinocervicothalamic, spinoreticular or spinomesencephalic tracts (see Fig. 4.5 and Table 4.3). Some of the tracts are bilateral, some are primarily contralateral. Some synapse in the grey matter several times en route. The key point is that there are multiple, bilaterally represented pathways for conveying noxious stimuli to the brain; this is clinically significant. For a spinal cord lesion to cause loss of nociception caudal to the lesion, there must be extensive destruction across the width of the cord to disrupt all the tracts. Therefore, clinically, loss of nociception is a poor prognostic sign in animals with severe spinal cord lesions (Fig. 6.4).
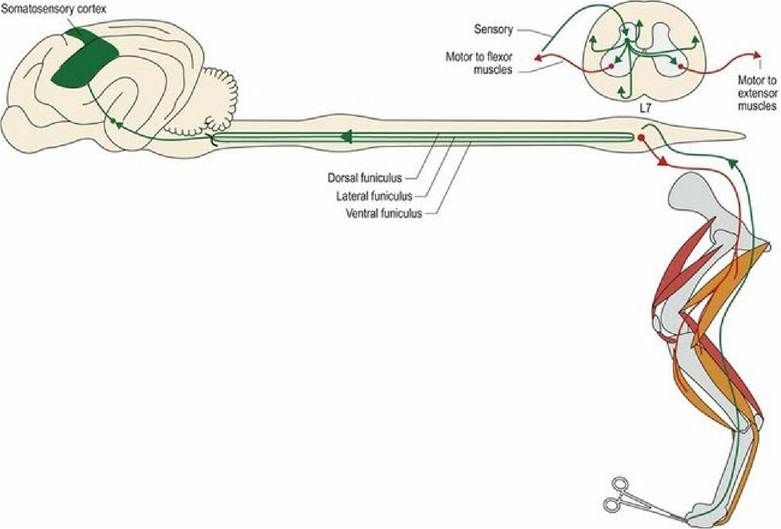
Fig. 6.4 Nociceptive input from the limbs and body induces local reflex functions and travels cranially in a number of different tracts, in the dorsal, lateral and ventral funiculi. Extensor muscles are red and flexor
muscles are orange.
In the normal animal, a noxious stimulus should cause a conscious response such that the animal whines, or looks at the site of the stimulus, as well as pulling the foot away reflexively (see Fig. 13.10). The withdrawal reflex will occur as long as the reflex wiring comprising the peripheral input, central spinal connections and the LMN to the flexor muscles is intact.
Thus an animal with a complete spinal cord transection at the thoracolumbar junction (cranial to the lumbar intumescence) should have an intact flexor withdrawal reflex in the pelvic limbs, but will not demonstrate signs of pain (i.e. conscious perception).The mechanisms of acupuncture
A-δ fibre input into the dorsal horn activates an enkephalinergic interneuron that inhibits C-fibre input into the same dorsal horn. Thus by stimulating the fast pain fibres, C-fibre input can be diminished. This is called the segmental effect as it acts within the stimulated spinal cord segment. Additionally, stimulation of ascending tracts triggers the release of a variety of neurotransmitters including β-endorphin, noradrenaline and serotonin from the brainstem, and oxytocin and adrenocorticotrophic hormone from the pituitary gland. Neurotransmitters can either act humerally or activate a descending analgesic system in the brain stem that decreases ascending pain signals. For example, nociceptive activation of the spinomesencephalic tract, may stimulate the periaqueductal grey matter of the midbrain, which then activates a descending inhibitory (analgesic) neural system. These are the heterosegmental effects as they are more generalised. Both the segmental and heterosegmental effects are most potent in the dorsal horn that is being stimulated (Fig. 6.5). Therefore pain relief is best achieved by needling as close as possible to the source of pain.
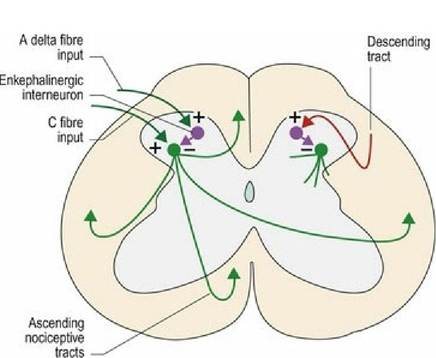
Fig. 6.5 Mechanisms underlying the segmental and heterosegmental effect of acupuncture.
Nociceptive input from head
All three branches of the trigeminal nerve convey nociceptive information into the brainstem where they synapse in the spinal nucleus of CN V in the medulla oblongata (see Chapter 10). Cranial nerves VII, IX and X, from the ear and oral cavity, also project to this nucleus (Fig. 6.6). Incoming fibres can then activate other brainstem nuclei for reflex function.
In the palpebral reflex, for example, a noxious stimulus travels via CN V to the spinal nucleus of V; interneurons project to CN VII facial nucleus stimulating eyelid closure. For conscious perception afferent axons from the nucleus of V, decussate and travel rostrally with medial lemniscus as the trigeminal lemniscus. They synapse in thalamus and are distributed to the somatosensory cortex. Thus the projection of nociception from the head is primarily contralateral, therefore unilateral lesions of forebrain will cause mainly contralateral hypoalgesia of the face.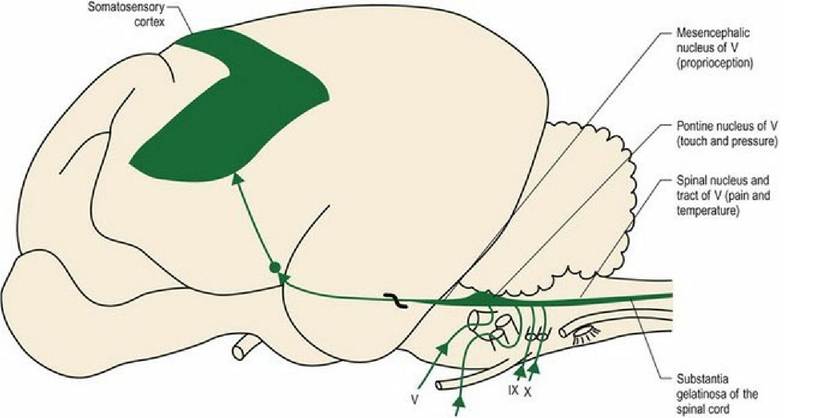
. vl1
Fig. 6.6 Nociceptive input from the head. The thick black wavy line represents decussation of the tract.
Nociceptive input from the viscera
Noxious visceral stimuli (see Chapter 12) are transmitted to the CNS via sympathetic nerves from the thoracic and abdominal viscera, and pudendal nerves from the pelvic viscera. Input from the respiratory tract and oesophagus is via the vagus nerve. Inputs via the sympathetic fibres will stimulate visceral reflexes such as accelerated heart rate and respiration (sympathetic response). Conversely, afferent fibres travelling in parasympathetic nerves may stimulate the parasympathetic system responses resulting in bradycardia and hypotension. Other inputs, for example distension of the gut wall, may stimulate reflex changes in gut activity, locally using the local autonomic plexi in the gut wall. Noxious stimuli from the viscera are also conveyed to the forebrain for conscious perception, but are often poorly localised (Fig. 6.7).
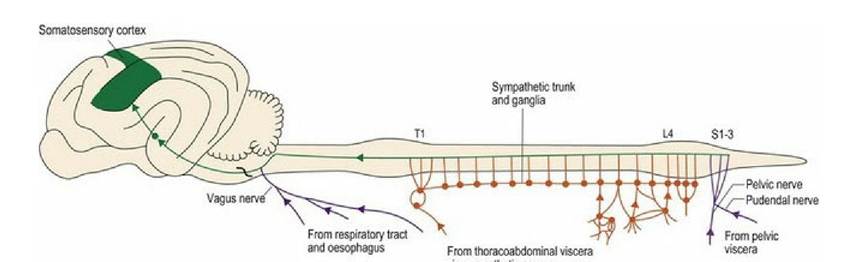
Fig. 6.7 Nociceptive input from the viscera uses nerves conveying sympathetic (orange) and parasympathetic (purple) fibres. Such input may induce autonomic reflex activity or stimulate conscious
awareness.