General proprioception
Proprioception (proprius - L = one’s own, capere - L = to receive) is the sense of the relative position of parts of the body. Proprioception indicates whether and how joints, muscles and tendons are moving based on input from muscle spindles, Golgi tendon organs and joint receptors.
It also indicates where various parts of the body are located in relation to each other. Tactile and mechanoreceptors, especially on the feet, contribute to proprioception, especially conscious proprioception, while hair cells in the vestibular apparatus of the inner ear supply information about head position and movement.Proprioceptive information from peripheral receptors travels via spinal nerves, the dorsal roots and spinal cord (dorsal and lateral funiculi) to the brain (Fig. 6.1). Proprioception of the head, its muscles and joints, uses mainly cranial nerves VIII and V to reach the appropriate brainstem nuclei. The information that terminates in the somatosensory cortex of contralateral cerebrum is used in conscious proprioception while that terminating in the ipsilateral cerebellum is used in subconscious proprioception. Conscious proprioception is the conscious awareness of body position and movement of body parts. It enables the cerebral cortex to plan and refine voluntary, learned movements. Subconscious proprioception is based around stretch and tension of muscles, tendons and ligaments, at rest and during movement, and spatial orientation of the body. The cerebellum needs this information to coordinate posture and locomotion. The vestibular system provides proprioceptive information (conscious and subconscious) about head position and movement. This input is fundamental for setting the balance and posture of the whole animal.
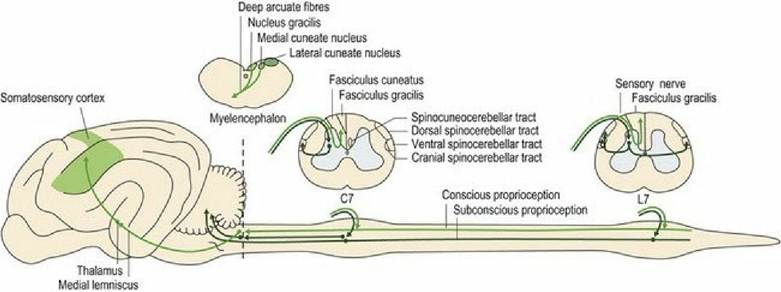
f⅛∙ 6∙1 Proprioceptive tracts to the brain from the body.
C7 = 7th cervical spinal cord segment, L7 = 7th lumbar spinal cord segment.Subconscious proprioception from the head (vestibular input), body and limbs (spinal input) is essential for normal posture and gait. Subconscious proprioceptive deficits result in the clinical sign of ataxia. Ataxia can be defined functionally as incoordinated movement. It presents as changes in the rate, range and force of movement. Ataxia does not mean that the animal is paretic (weak), however, cases of spinal cord disease may often present with both ataxia and UMN paresis due to compression of general proprioceptive and UMN tracts.
Both conscious and subconscious proprioceptive systems are essential for normal posture and gait and it can be difficult to rigorously separate conscious from subconscious proprioceptive deficits when presented with an ataxic patient. However in dogs and cats, subconscious proprioceptive deficits usually perturb posture and gait more than conscious proprioceptive deficits. This is evidenced by observing the effect of lesions located at the termination of subconscious versus conscious proprioceptive pathways. For example, ataxia is more pronounced with lesions in the cerebellum than lesions affecting the somatosensory cortex in the forebrain. Subconscious proprioceptive deficits due to cerebellar lesions may present with truncal sway, base wide or narrow posture and limb movement, and delays in initiating or terminating movement. In dogs and cats with lesions affecting the somatosensory cortex where conscious proprioceptive pathways terminate, only
mild stumbling, or a tendency to stand on the dorsum of the paw, may be observed. Gait and posture may otherwise be reasonably normal.
Conscious proprioception
Key points
■ Conscious proprioception is the information about the position of the head, body and limbs that is received in the contralateral somatosensory cortex of the cerebrum and is used particularly for executing voluntary, skilled movement.
■ Information from the body and limbs is transmitted via the spinal nerves and spinal cord.
■ Information from the head is transmitted via CN V (somatic sensation and muscle proprioception) and CN VIII (head position and movement).
■ Conscious proprioception utilises sensory inputs from touch, pressure, muscle and joint receptors. Hair cells of the vestibular apparatus transmit information about head position and movement.
Non-painful, conscious sensations can broadly be divided into two categories that share functional and anatomical features. One group comprises touch and pressure, and the other is composed of joint and muscle/tendon proprioception. Both are principally transmitted to the cerebral cortex via the dorsal column- medial lemniscal system involving primarily the dorsal funiculus of the spinal cord with some transmission in the lateral funiculus (see Fig. 4.5). The dorsal funiculus is also known as the dorsal column and it continues as the medial lemniscus in the brainstem.
Axons conveying this information enter the dorsal funiculus and join the fasciculus cuneatus (from the body cranial to T8) or fasciculus gracilis (caudal to T8). Fibres in the fasciculus cuneatus convey proprioceptive and discriminative touch. They synapse in the medial cuneate nucleus located in the caudal medulla oblongata (see Figs. A26-30). Only about one-quarter of the fasciculus gracilis fibres synapse in the nucleus gracilis in the medulla oblongata. Thus the dorsal funiculus is relatively unimportant in transmitting proprioception from the pelvic limbs in quadrupeds; it is important in humans. The majority of fibres entering the fasciculus gracilis leave it in the cranial lumbar region to synapse in the grey matter on interneurons, or relay neurons of other pathways, or even LMNs. Fibres arising from synapses at the base of the dorsal horn (e.g. the nucleus thoracicus (see Table 4.2) may continue cranially in the dorsal spinocerebellar tract (with collateral fibres travelling in the adjacent spinomedullary tract) to synapse near the nucleus gracilis (in nucleus Z) (Figs.
A24, A27, A28). Post-synaptic fibres from these proprioceptive relay nuclei (gracile, medial cuneate and Z) decussate in the deep arcuate fibres of the medulla oblongata and continue rostrally in the medial lemniscus to the thalamus where they synapse. They are projected to the somatosensory cortex of the cerebrum via the internal capsule.The role of the lateral funiculus in conveying pelvic limb proprioception to the cerebrum may be one explanation why cervical spinal cord compression can produce particularly marked pelvic limb ataxia. This is illustrated in Fig. 6.3 on tWobbler' horses and dogs (see accompanying text).
The dorsal column-medial lemniscal system is highly organised topographically and is quite specific with respect to the transmission of sensory and discriminatory information. In other words, there is little convergence (input) of other pathways onto the relay nuclei; this enables the animal to localise precisely a stimulus from the skin, e.g. the horse biting at a fly on its body. Similarly, the somatosensory cortex is somatopically arranged and the size of each area reflects the density of innervation (see Fig. 4.14).
The dorsal funiculus comprises a greater percentage of white matter in carnivores and primates than in other animals; this reflects higher sensory discrimination from the digits, especially compared with ungulates.
There are two kinds of proprioceptive input from the head - that associated with head position and movement (see Chapter 8) and proprioception due to receptors associated with muscles of mastication, facial expression and eye movement. Muscle proprioception is also conveyed by a three-neuron system via all three branches of CN V (trigeminal nerve). The trigeminal ganglion at the base of the neurocranium is the equivalent of a spinal ganglion, containing somatosensory neuronal cell bodies. However, uniquely, the cell bodies of the primary afferent neurons concerned with muscle proprioception are found in the CNS, in the mesencephalic nucleus of V.
The second stage fibres decussate, travel rostrally in the trigeminal lemniscus to the thalamus and on to the somatosensory cortex. The proprioceptive fibres in the mesencephalic nucleus of CN V also connect to other cranial nerve motor nuclei for reflex function. Sensory fibres from tongue musculature (intrinsic and extrinsic) have their cell bodies located in the trigeminal, C1 and distal vagal ganglia.Subconscious proprioception
Key points
■ Subconscious proprioception is information about the position of the head, body and limbs that is received in the cerebellum and is used for coordinating posture, locomotion and semi-automatic movement.
■ It is transmitted from receptors such as muscle spindles and Golgi tendon organs of the body and head musculature, or the vestibular apparatus of the head, to the ipsilateral cerebellum.
Subconscious proprioceptive information is received by the ipsilateral cerebellum and is used for coordinating posture and movement. Receptors involved in subconscious proprioception are primarily muscle spindles and Golgi tendon organs, and the vestibular system of the head. Conscious awareness of this type of proprioceptive information is not required for normal function. However, collateral axons also project to the cerebrum contributing to conscious proprioception and awareness of movement (kinaesthesia).
Subconscious proprioceptive pathways inform the cerebellum about location, status (tension and length) and movement of the skeletal musculature throughout the body, limbs and head. The cerebellum also receives copies of planned motor activity from the motor cortex and UMN centres. Thus it compares incoming proprioceptive sensory information with the motor plans and generates an output back to motor centres to adjust and perfect the muscular activity (see Chapter 7).
Spinocerebellar pathways comprise two neurons, compared with the three-neuron pathway involved in conscious proprioception. The first neuron originates at the peripheral receptor and has its cell body in the spinal ganglia.
It synapses in the base of the dorsal horn and post-synaptic fibres project via the lateral funiculus to the cerebellum. The exception to this is the spinocuneocerebellar tract, fibres of which pass through the dorsal horn into the fasciculus cuneatus and synapse in the lateral cuneate nucleus of the medulla oblongata and post-synaptic neurons project to the cerebellum. Note that the lateral cuneate nucleus and superficial arcuate fibres are associated with subconscious proprioception, while the medial cuneate nucleus and deep arcuate fibres function in conscious proprioception.Information from the caudal half of the body travels cranially in the dorsal and ventral spinocerebellar tracts. The dorsal spinocerebellar tract (DSCT) arises from the nucleus thoracicus at the base of the dorsal horn (see Fig. 4.5 and Table. 4.2) located from C8-L4. Fibres enter the lateral funiculus, and travel cranially to the medulla oblongata, into the caudal cerebellar peduncle and terminate in the ipsilateral vermis and paravermis (see Chapter 7). Therefore the DSCT serves both to project to the cerebellum as well as to the sensory area of the cerebral cortex.
The ventral spinocerebellar tract (VSCT) also arises from the nucleus thoracicus, but the fibres decussate to contralateral lateral funiculus and course cranially to the rostral cerebellar peduncle. Here fibres decussate again to end up in the cerebellum ipsilateral to the side of the stimulus.
Information from the cranial half of the body T8-C1 is conveyed cranially by three routes. The spinocuneocerebellar tract travels cranially in the fasciculus cuneatus to synapse in the medulla oblongata (lateral cuneate nucleus), then via the caudal cerebellar peduncle to the ipsilateral cerebellum. The cranial spinocerebellar tract (CSCT) arises from neurons at the base of the dorsal horn, and travels in the ipsilateral lateral funiculus entering the cerebellum via both the rostral and caudal cerebellar peduncles. The cervicospinocerebellar tract relays information concerned with proprioception of the neck which is essential for whole body postural control (see Chapter 8).
There is variation in these tracts between the species, reflecting the species’ specific movement capabilities. The dorsal and ventral spinocerebellar tracts are well developed in ungulates. Cats also have the cranial spinocerebellar tracts and spinocuneocerebellar tracts, which enables them to have highly coordinated function of the thoracic limb and forepaw.
Subconscious proprioception from the body, limbs and head is represented ipsilaterally in the cerebellum. Tracts that decussate en route to the cerebellum, like the ventral spinocerebellar tract, decussate again once inside the cerebellum. Fibres involved in vestibular-cerebellar connections always remain ipsilateral. Conversely, input to the cerebellum from the motor planning and UMN centres is represented contralaterally, but cerebellar output to these centres also decussates, thus influencing the original side.
In humans, kinaesthesia is the conscious awareness of body position and movement. We can assume it occurs in animals, such that an animal is aware of its body position and movement, but not of the specific action of each muscle. Proprioceptive information used in kinaesthesia arises from collaterals of subconscious proprioceptive pathways. Collateral information from the caudal half of the body travelling in the dorsal spinocerebellar tract, synapses in the medulla oblongata (nucleus Z). Similarly, collateral information from the cranial half of the body travelling in the spinocuneocerebellar pathway, synapses in the medulla oblongata near the lateral cuneate nucleus (nucleus X). From nucleus Z and nucleus X, fibres join the medial lemniscus for projection to the contralateral thalamus and then to the cerebral cortex.
Conscious and subconscious proprioception in posture and gait
Key points
■ Conscious proprioceptive deficits are typified by the animal bearing weight on an abnormal part of the foot - e.g. on the dorsum of the paw.
■ Subconscious proprioceptive deficits are typified by an abnormal position of the limbs (props) with respect to the centre of gravity, at rest and during locomotion.
A key concept in clinical neurology is that similar neurological deficits will be produced regardless of where a lesion is located along a neural pathway - origin, midway or the termination. Applying this concept can help illustrate the role of conscious versus subconscious proprioception in dogs and cats, by considering what proprioceptive deficits may be present in an animal with a lesion affecting the termination of the conscious versus subconscious proprioceptive pathway. What happens when proprioceptive information is not received at the somatosensory cortex or at the cerebellum? With somatosensory cortex lesions, the gait is often quite coordinated with minimal ataxia. The limbs are placed in a weight-bearing position under the body at rest and during movement. However the animal may stumble and come to rest standing on the dorsum of the paw (‘knuckling’) (Fig. 6.2); these signs will be contralateral to the side of the lesion due to decussation of the sensory pathway in the medulla oblongata (see Fig. 4.10). Deficits reflect loss of input especially from tactile and joint angle receptors. Conversely, the animal with cerebellar lesions may show pronounced subconscious proprioceptive deficits in which the limbs will not be placed in a good weight-bearing position under the centre of gravity. At rest the animal may have a base wide or narrow stance, and during locomotion, the limbs may be abducted, adducted or cross over. These signs reflect failure to receive and process input, especially from muscle spindles of the proximal limb in particular from the extrinsic limb muscles (those that attach the limb to the trunk). Extrinsic muscles provide information about the angle the limb makes with the body. Conversely, a spinal cord lesion may compromise both conscious and subconscious proprioceptive pathways: thus the animal is ataxic and has deficits in the paw position response and may stand knuckled.
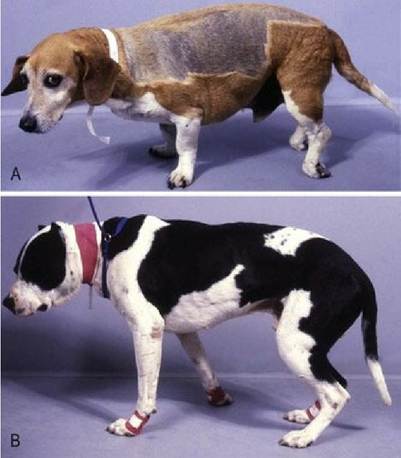
Fig. 6.2 (A) Dog with a lumbar spinal cord lesion resulting in abnormal paw position sense and ‘knuckling’; this sign is characteristic of involvement of the conscious proprioceptive pathways. (B) The limbs are not under the centre of gravity in this dog; this sign is characteristic of dysfunction of the subconscious proprioceptive pathways.
Clinically, tests that specifically assess conscious proprioception (along with motor function) stimulate the tactile receptors in the foot. Thus the paw position response (‘knuckling’) and hopping are useful tests. Hopping also stimulates muscle spindle receptors throughout the limb, while the paw position response test stimulates those in distal extensor/flexor muscles in the foot. For subconscious proprioceptive testing, tests that change muscle and tendon inputs by altering the relative position of the limbs under the centre of gravity are useful and include hopping, and reflex stepping (see Chapter 13).
Note: to clinically make an absolute distinction between the two types of proprioception may not be possible and, for spinal cases, is not necessary. Also note that an animal must have a functioning motor system to demonstrate normal proprioception. For example, it must be able to move its foot after the paw is turned over, or lift the limb and place it in a new weight-bearing position during the hopping test. If the animal has significant motor dysfunction (UMN or LMN) it may not be able to perform the test even if the proprioceptive tracts are intact. Thus lesions that disrupt motor systems may lead to apparent proprioceptive dysfunction. Differentiating between proprioceptive versus motor dysfunction is discussed in Chapter 13.
Spinal lesions can compromise the proprioceptive input from the limbs and also from the trunk. The cerebellum needs proprioceptive input from the axial muscles to coordinate contraction of spinal agonists and antagonists. Loss of subconscious proprioceptive input from the trunk explains the difference in the clinical signs observed in a cranial thoracic versus mid-lumbar lesion. In both cases, the animal will have deficits in pelvic limb function. However, with the cranial thoracic cord lesions there may also be truncal ataxia, and the body may list to the side. Such lesions may also affect motor systems resulting in paresis of the trunk muscles.
The effect of spinal cord lesions on general proprioception is illustrated in horses and dogs with ‘wobbler syndrome’ caused by compressive lesions in the cervical spinal cord. Characteristically, gait deficits and ataxia are more marked in the pelvic limbs, compared with the thoracic limbs. In part this is because the dorsal spinocerebellar tracts from the pelvic limbs are usually more severely compressed than those tracts in the dorsal funiculus conveying proprioception from the thoracic limbs. Loss of general proprioceptive input results in typical, wobbly gait. With loss of conscious proprioception, the dog may stand on the dorsum of the paw (Fig. 6.3A) or the horse, when turned in a tight circle, may spin on the hoof rather than stepping around in a circle. Wobbler animals also exhibit ataxia, failing to keep the limbs in a good weight-bearing position under the centre of gravity either at rest or during locomotion (Fig. 6.3B). This sign is typical of subconscious proprioceptive deficits. These cervical lesions often affect UMN tracts also, resulting in paresis. Caudal cervical lesions can compromise LMN innervation to the shoulder muscles, resulting in a shortened stride in the thoracic limbs due to reduced shoulder extension. It may also cause prominent, specific atrophy of the affected muscles; this is called neurogenic atrophy as it arises due to loss of peripheral motor neuron (LMN) innervation (Chapter 5).
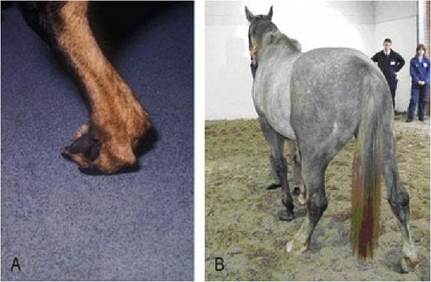
Fig. 6.3 Wobbler animals with proprioceptive deficits: (A) dog with delayed paw position response and in (B) the horse that would stand base wide for prolonged periods.