Oogenesis
In the fetus, primordial germ cells migrate from the yolk sac to the developing ovaries, where a single layer of follicular cells surrounds a germ cell destined to become an ovum.
The central germ cell (now termed an oogonium) enlarges and begins meiosis. (Recall that meiosis entails two cell divisions during which the diploid number of chromosomes is reduced by half to the haploid number.) The oogonium does not complete meiosis; it stops in the first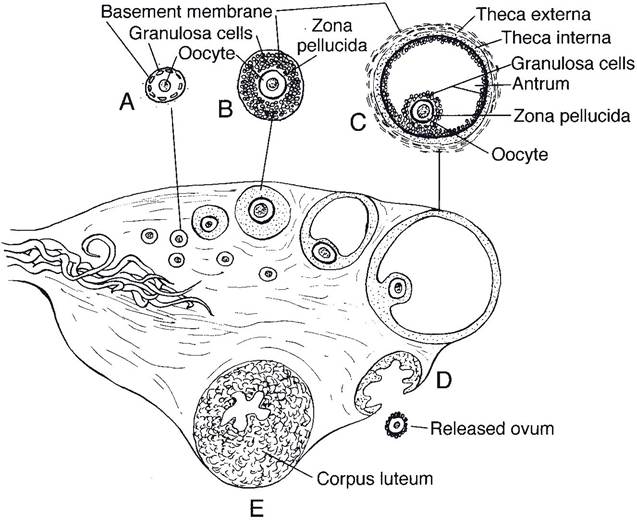
Figure 27-1. Sagittal section of an ovary showing the origin, growth, and ovulation of a follicle and a corpus luteum developing at the site of a follicle that has ovulated. A, Primary follicle; B, growing follicle; C, Graafian or tertiary follicle; D, ovulation; E, corpus luteum. (Reprinted with permission of Wiley-Blackwell from Reece W.O. Physiology of Domestic Animals. 2nd ed. Baltimore: Williams & Wilkins, 1997.)
prophase before the first division. At this stage, the developing ovum is a primary oocyte, and the combination of a primary oocyte and its surrounding cuboidal follicular cell (granulosa cell) layer is a primary follicle (Fig. 27-1). At birth, the ovaries of most domestic species contain hundreds of thousands of primary follicles waiting to continue their development. What determines which of the thousands of primary follicles is selected to develop further during a specific estrous cycle is unknown.
in contrast to spermatogenesis, which produces four spermatozoa from each primary germ cell, the maturation of the primary oocyte results in only one mature ovum and three rudimentary cells, called polar bodies. In most animals, the first of the two meiotic divisions is complete, resulting in the formation of the first polar body, before or immediately after ovulation (the discharge of an oocyte from a follicle).
Secondary Follicles
in all animals, multiple primary follicles typically begin further development during a single estrous cycle. in monotocous animals (animals not bearing litters and normally having only one offspring per gestation, such as the mare and cow), one follicle usually develops more rapidly than others, and only one ovum is released at ovulation. The rest of the developing follicles regress and form atretic follicles. Polytocous animals, such as carnivores and swine, which normally produce two or more offspring per gestation, usually have several follicles that develop and ovulate at approximately the same time. The ova may all come from one ovary, or some may come from each ovary.
The further development of primary follicles includes enlargement of the oocyte and replication of the surrounding follicular cells. The replicating follicular cells become several layers thick, and this surrounding group of cells is a granulosa. The granulosa cells secrete glycoproteins that cross-link to form a protective shell, the zona pellucida, around the oocyte (Fig. 27-1). Cytoplasmic processes of granulosa cells penetrate the zona to permit communication and exchange between them and the oocyte. The initial development to this point is independent of hormonal stimulation by gonadotrophins (follicle-stimulating hormone [FSH] and luteinizing hormone [LH]).
The developing follicle is termed a secondary follicle when the oocyte has enlarged and is surrounded by a developing granulosa. A theca, consisting of layers of cells immediately surrounding the granulosa, also first develops late during the secondary follicle stage.
Hormones and Follicular Development
Gonadotrophin-releasing hormone (GnRH) is released from the hypothalamus to promote the release of both FsH and LH from the adenohypophysis. The release of GnRH can be modulated by steroid (estradiol and progesterone) and peptide (inhibin) hormones from the ovary, but its basal release is determined by neural inputs to the hypothalamus.
The basal release of GnRH is assumed to be pulsatile, for this is the type of release seen in ovariectomized animals. The pulsatile nature is physiologically important, because continuous infusions of GnRH do not result in the continuous release of FsH and LH.The granulosa and theca of secondary follicles develop cellular receptors for FsH and LH, respectively, and become responsive to these hormones. From this point, the coordinated effects of FsH and LH are both needed for normal follicular development. Under the influence of LH, thecal cells proliferate and produce androgens (androstenedione and testosterone) that diffuse into the granulosa. FsH promotes further granulosa cell proliferation, the development of cellular enzymes necessary for the conversion of androgens to estrogens (estradiol), and the secretion of several other paracrine agents necessary for follicular development.
The cellular secretions accumulate among the granulosa cells, and ultimately a fluid-fllled cavity (antrum) can be identified. The developing follicles are tertiary follicles (also known as vesicular or Graafian follicles) when an antrum can be identified among the granulose cells (Fig. 27-1). Theca surrounding tertiary follicles have two layers, the theca externa and theca interna (Fig. 27-1). The internal layer is highly vascular and contains thecal cells with cellular characteristics of steroid-producing cells. The theca externa primary consists of connective tissue.
The estrogen produced by the granulosa cells acts as a paracrine agent on the developing follicle and also enters the systemic circulation to affect other sites throughout the body (Fig. 27-2). Locally, the estrogen acts on granulosa cells to increase FsH and LH receptors, and, together with these gonadotrophins, it promotes further granulosa cell replication, growth, and secretion. The overall effect is that locally produced estrogens promote the development of the follicle from which they are being produced.
This is characterized as a local positive feedback effect of the estrogens. This positive feedback effect is one factor in the selection process that determines which of the developing follicles will ultimately produce the ovum and ovulate. A second factor is that circulating estrogens have a negative feedback effect on FSH secretion from the adenohypophysis. The decrease in FsH during this period contributes to the atresia of more slowly developing follicles. The selection process also involves another follicular hormone, inhibin, discussed later.Estrogens from developing follicles are also necessary to prepare the follicles and the hypo- thalamic-adenohypophyseal axis for ovulation. Within the ovary, estrogens promote an increase in LH receptors in thecal cells so that these cells increase their production of androgens and appropriately respond to LH at the time of ovulation. Circulating estrogens promote an increase in LH within the adenohypophysis and condition the hypothalamic-adenohypophyseal axis so that the short-term large LH release (termed LH surge) necessary for ovulation can be delivered (Fig. 27-2).
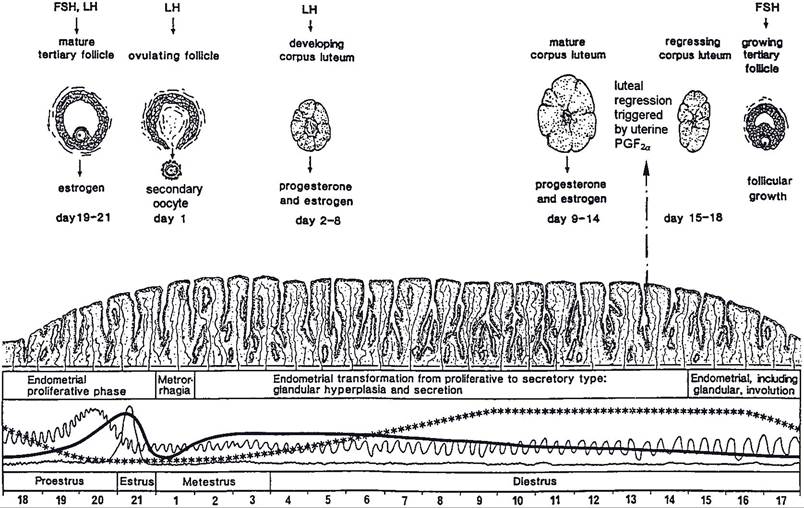
Figure 27-2. Ovarian, uterine, and hormonal changes during the estrous cycle of the cow. The scale indicates days 1 to 21 of the cycle. Relative blood levels of progesterone (***), estrogen(-), FSH(^A^∣∩), and LH (—)are shown. (Reprinted with permission of Wiley-Blackwell from Dellmann H.D. and Eurell J. Textbook of Veterinary Histology. 5th ed. Baltimore: Lippincott Williams & Wilkins, 1998.)
Non-litter-bearing animals typically have one or two follicles per estrous cycle that develop faster and grow larger than the rest. These are dominant follicles. In primates there is typically only one dominant follicle per estrous cycle, and it provides the ovum. The development of the dominant follicle is accelerated after the corpus luteum (discussed later) from the previous estrous cycle has regressed (Iuteolysis).
The phase of the estrous cycle in primates during which there is no corpus luteum and the dominant follicle is developing is the follicular phase of the estrous cycle. The luteal phase is the part of the cycle during which a corpus luteum is intact and secreting progesterone. Formation of a corpus luteum and its functions are discussed later in this chapter.in domestic animals that typically have only one or two offspring per pregnancy, large dominant follicles may develop while a corpus luteum remains intact. These dominant follicles may or may not ovulate. Mature follicles that do not ovulate undergo atresia if a corpus luteum remains intact. At that point, another dominant follicle begins to develop rapidly so that ovulation can occur soon after luteolysis. Because dominant follicles may develop while a corpus luteum remains intact, the estrous cycles of large domestic animals are considered to have overlapping follicular and luteal phases.
Inhibins are peptide hormones secreted by granulosa cells of developing follicles. Circulating levels of inhibins increase with follicular development, and inhibins have a negative feedback effect on FSH release from the adenohypophysis. By this means, a developing dominant follicle can suppress the development of competing follicles in non-litter-bearing animals. in litter-bearing animals, the combined negative feedback effect of inhibins from multiple follicles can suppress other follicles to prevent litter sizes from becoming inappropriately large. inhibins from developing follicles apparently do not suppress LH secretion necessary for ovulation.