OVARIAN FOLLICLE ACTIVITY
1. How do growing follicles become Graafian follicles?
2. What part of the Graafian follicle secretes androgens? Do androgens persist as androgens?
3. What hormones cause the formation of a fluid-filled space called an antrum?
4.
What functions are served by the preovulatory (24 hours) surge of LH?5. Do all animals ovulate before the end of estrus? What is the difference between spontaneous and reflex ovulation?
6. Does ovulation occur in all developing follicles? Do follicles continue to grow and develop during all phases of the ovarian cycle? What must be a characteristic of follicles for them to ovulate?
7. What changes are involved in the formation of the corpus luteum? How is the corpus luteum maintained?
8. What is the natural luteolytic substance that causes regression of the corpus luteum? Does acute regression of the corpus luteum occur in the bitch and queen?
9. Note the unique delivery system of the natural luteolytic substance.
0. What is a persistent corpus luteum and what is its most probable cause?
When reproductive cycling begins, select follicles within the ovary are influenced by hormones and proceed through growth and maturity, followed by ovulation, corpus luteum development, and its regression. These changes reoccur for other follicles at intervals characteristic for a species.
Follicular Growth
Puberty is defined as the beginning of reproductive life, which in the female is usually marked by the beginning of ovarian activity. The formation of Graafian follicles from growing follicles is hormone dependent and begins at puberty when tonic levels of LH and FSH begin to rise and fall with each estrous cycle. Interstitial cells begin to surround the basement membrane of the granulosa cells to form the theca, which differentiates into a theca interna and externa. As the thecal cells are formed around the follicle, a capillary bed develops among them.
These thecal capillaries increase in size and are concentrated in the theca interna close to the basement membrane that separates the theca interna cells from the granulosa cells (Figure 15-15). LH receptors form on the cells of the theca interna, and receptors for FSH and estrogen form on the granulosa cells.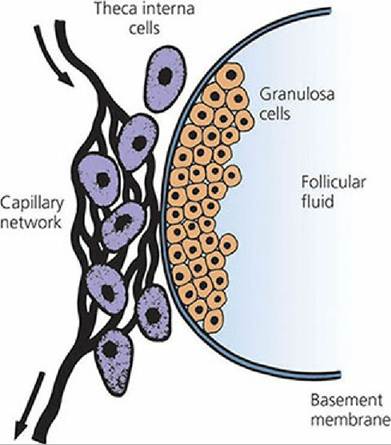
■ FIGURE 15-15 Formation of a Graafian follicle from a growing follicle. Wall structure. The theca interna cells are well supplied with blood. The basement membrane deprives granulosa cells of blood supply. (From Baird DT. Reproductive hormones. In: Austin CR, Short RV, eds. Reproduction in Mammals, Book 3. Cambridge, England: Cambridge University Press, 1972. Reprinted by permission of Cambridge University Press.)
During the hormone-dependent stage, under the influence of LH, androgens are produced by cells of the theca interna. The androgens diffuse from the theca interna to the granulosa cells. Under the influence of FSH, the granulosa cells convert the androgens to estrogens. The estrogens produced cause growth and division of the granulosa cells and, together with FSH, cause the granulosa cells to produce secretions that effect separation of the granulosa cells and formation of a space filled with fluid (liquor folliculi), called an antrum (see Figure 15-15). FSH also stimulates the formation of LH receptors on the granulosa cells. A surge of LH output (preovulatory surge) occurs about 24 hours before ovulation. In addition to its role in ovulation and formation of a corpus luteum, the LH surge causes a reduction in the number of FSH receptors on granulosa cells, so that the output of estrogen by the granulosa cells decreases.
Ovulation
When the oocyte is released into the abdomen from its protruding follicle, it is covered by those granulosa cells that immediately surrounded it just before ovulation; these are known as the corona radiata. The oocyte and granulosa cells are evacuated with an enveloping viscous (gelatinous) follicular fluid.
At ovulation, the oocyte, together with its surrounding cells and gelatinous mass, is swept into the uterine tubes by motility of the fimbriae. The relationship of ovulation to estrus for domestic animals and other factors involved in female reproduction are given in Table 15-1.| TABLE 15-1 FACTORS RELATED TO FEMALE REPRODUCTION | |||||
| ANIMAL | ONSET OF PUBERTY (mo) | AGE FIRST SERVICE (AVERAGE) | LENGTH OF ESTROUS CYCLE (D) | LENGTH OF Estrus | GESTATION PERIOD (D) |
| Mare | 18 (10-24) | 2-3 yr | 21 (19-21) | 5 d (4.5-7.5 d) | 336 (323-341) |
| Cow | 4-24 | 14-22 mo | 21 (18-24) | 18 h (12-28 h) | 282 (274-291) |
| Ewe | 4-12 (first fall) | 12-18 mo | 16-1/2 (14-20) | 24-48 h | 150 (140-160) |
| Sow | 3-7 | 8-10 mo | 21 (18-24) | 2 d (1-5 d) | 114 (110-116) |
| Bitch | 6-24 | 12-18 mo | 6-12 mo | 9 d (5-19 d) | 63 (60-65) |
| TIME OF OVULATION | OPTIMUM TIME FOR SERVICE | ADVISABLE TIME TO BREED AFTER.PARTURITION | |||
| Mare | 1-2 d before end of estrus | 3-4 d before end of estrus or 2nd or 3rd d of.estrus | About 25-35 d or second estrus about 9 d or first estrus only if normal in every way | ||
| Cow | 10-15 h after end of estrus | Just before middle of estrus to end of estrus | 60-90 d | ||
| Ewe | 12-24 h before end of estrus | 18-24 h after onset of estrus | Usually the next fall | ||
| Sow | 30-36 h after onset of estrus | 12-30 h after onset of estrus | First estrus 3-9 d after weaning pigs | ||
| Bitch | 1-2 d after onset of true estrus | 2-3 d after onset of estrus or 10 14 d after onset of proestrous bleeding | Usually first estrus or 2-3 mo after weaning pups | ||
| Data from Frandson RD, Spurgeon TL. Anatomy and Physiology of Farm Animals. 5th edn. Philadelphia. PA: Lea & Febiger, 1992. | |||||
Ovulation is spontaneous (no stimulation needed) in all of the domestic species except the cat. The cat and other nonspontaneous ovulators (e.g., mink, rabbit, ferret) are reflex ovulators, in that coitus is required for ovulation to occur. Coital contact apparently brings forth an LH surge.
The selection of follicles for ovulation seems to occur primarily by chance. It is usually associated with the largest actively growing follicles present when the previous CL regressed (i.e., when progesterone decreased and FSH and LH output began to increase). Follicles continue to grow and develop during all phases of the ovarian cycle, with some impairment during the luteal phase, and the LH surge is necessary for ovulation to occur. Follicles close to full development, but without adequate LH receptors, do not ovulate in response to the LH surge and become atretic.
Corpus Luteum Formation and Regression
Formation of the CL involves luteinization of the granulosa, by which the granulosa is converted from estrogen secretion to progesterone secretion (LH receptors on the granulosa cells were previously induced by FSH). The process is initiated by the preovulatory LH surge. The cavity of the ruptured follicle and the fibrin clot within serve as the framework on which the granulosa cells develop. Blood vessels from the theca externa invade the developing CL and it becomes vascularized. Maintenance of the CL is provided for by LH derived from the LH surge and by the basal circulating levels of LH. In the sheep, prolactin, a gonadotropic hormone for some species, is required to maintain the CL, in addition to LH.
The uterus (endometrium) plays a major role in controlling the lifespan of the CL in nonpregnant mares, cows, sows, ewes, and does (goats) but is not active in CL regression in the bitch (dog) and queen (cat). PGF2α is released by the nonpregnant uterus about 14 days after ovulation and is considered to be the natural luteolytic substance (causes regression of the CL).
The venous return of uterine blood to the right heart and from there to the lung before transport of arterial blood to the ovary results in inactivation by the vascular endothelium of about 90% of PGF2α. To ensure that enough PGF2α is delivered directly to the ovary for luteolysis, the anatomic arrangement of the ovarian vein, which is returning blood from both the ovary and uterus, and the ovarian artery is such that PGF2α can diffuse from the vein to the artery and ovarian perfusion of PGF2α can occur before circulation through the lungs (Figures 15-11 and 15-16). For PGF2α to be effective when it enters the general circulation, it must either be secreted by the uterus in larger amounts or be more resistant to degradation in the lungs, or both. Survival of PGF2α in the general circulation is more important in the sow and mare.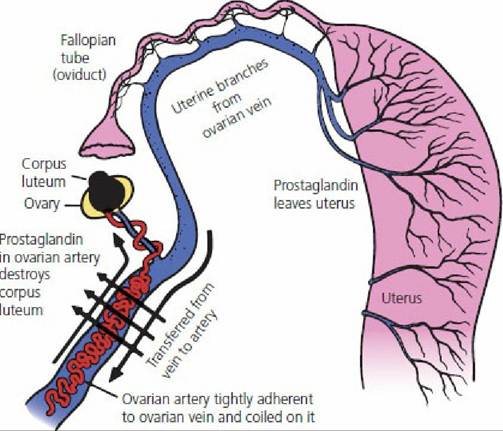
■ FIGURE 15-16 Postulated route by which prostaglandin secreted by the progesterone-primed uterus can enter the ovarian artery and destroy the corpus luteum in the ewe, and possibly other species. (Adapted from Short RV. Role of hormones in sex cycles. In: Austin CR, Short RV, eds. Reproduction in Mammals, Book 3. Cambridge, England: Cambridge University Press, 1972.)
The reason for final regression of the CL in the bitch and queen (bitch, 75 days; queen, 35 days) is not known. An acute lytic process does not occur.
Persistent Corpus Luteum
Prolongation of the luteal phase beyond 14 days to perhaps 1 to 5 months is known as persistent corpus luteum. The presence of a persistent CL prevents a return to the follicular phase and its next ovulation. The immediate reason for persistent CL is the failure of the endometrium to synthesize PGF2α. Often the failure is caused by an acute or chronic endometrial inflammation.
Summary of Ovarian Cycle Events
Events in the ovary associated with a cycle of hormone changes can be summarized as follows:
1.
After regression of the CL (luteolysis caused by PGF2α), FSH and LH secretion increases (because of a decrease in the concentration of progesterone).2. LH stimulates secretion of androgens by the theca interna cells, which diffuse into the granulosa cells.
3. FSH stimulates conversion of androgen to estrogen by the granulosa cells, and the estrogen concentration gradually increases.
4. FSH stimulates the formation of LH receptors on the granulosa cells.
5. Estrogen-rich fluid formed by the granulosa cells separates the granulosa cells and forms a pocket known as an antrum.
6. The gradually increasing estrogen concentration causes a preovulatory surge of LH release.
7. The LH surge promotes the maturation of oocytes by resuming meiosis through the first polar body stage.
8. The LH surge promotes the intrafollicular production of prostaglandins A and E (PGA and PGE), associated with rupture of the follicle.
9. Concomitant with PGA and PGE production is the formation of multivesicular bodies (MVB), which form as out-pockets of the exposed theca externa.
0. MVBs seem to secrete proteolytic enzymes that digest ground substance cementing the theca externa fibroblasts, allowing escape of the oocyte (ovulation).
11. The LH surge causes reduction in the number of FSH receptors on the granulosa cells, so the rate of conversion of androgen to estrogen diminishes.
.2. LH attaches to granulosa cell LH receptors and begins the conversion of the granulosa from estrogen secretion in the follicular phase to progesterone secretion in the luteal phase.
.3. At some point in the latter stages of these events, ovulation occurs and,the cavity previously occupied by the mature follicle becomes a corpus luteum.
.4. The corpus luteum secretes progesterone, which causes a decrease in the output of FSH and LH by the anterior pituitary.
L5. The corpus luteum regresses and the output of progesterone begins to decrease.
.6. A decrease in the level of progesterone causes FSH and LH secretion to increase, and the cycle is repeated.
The ovarian events are illustrated in Figure 15-17.
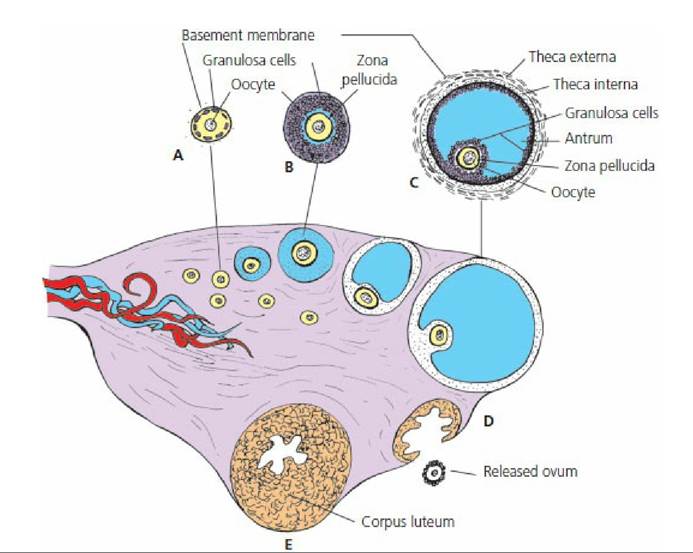
■ FIGURE 15-17 Sagittal section of an ovary. A. Primary follicle. B. Growing follicle. C. Graafian follicle. D. Ruptured follicle. E. Corpus luteum. This schematic representation shows in sequence the origin, growth, and rupture of a Graafian follicle and a corpus luteum that develops from the remains of the ruptured follicle.
■