Photoreception and Transduction of Light Occur in the Rods and Cones
The anatomical structures of the rod and cone photoreceptors are similar, but there are some important differences. Because they are neurons, the rods and cones are primary receptors.
Both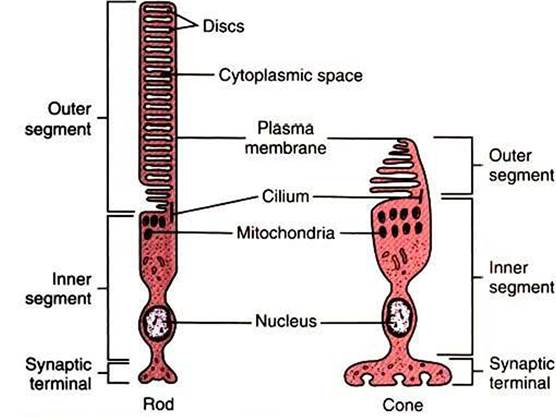
FIGURE 14-6 The two types of photoreceptors, rods and cones, have characteristic structures. Both rod cells and cone cells are differentiated into inner and outer segments connected by a cilium.The inner segments of both cell types contain the nucleus and most of the cell's biosynthetic machinery and are continuous with the synaptic terminals. The membranous discs in the outer segment contain the light-transducing photopigment.The discs in the outer segments of rod cells are separated from the plasma membrane, whereas the discs of cone cells are not. (Modified from O'Brien DF:The chemistry of vision, Science 218:961, 1982; from Kandel ER, Schwartz JH: Principles of neural science, ed 2, NewYork, 1985, Elsevier Science Publishing.)
cell types are divided into three parts: a synaptic terminal, an inner segment, and an outer segment (Figure 14-6). The photoreceptor synaptic terminal synapses with the bipolar cells. The inner segment includes the nucleus, mitochondria, and other cytoplasmic structures. The inner and outer segments are connected by a microtubule-containing cilium. The outer portions are specialized for photoreception. They contain an elaborate array of stacked membranous discs whose membranes contain visual photopigments. Visual photopigment is a light-sensitive molecular complex that initiates a biochemical chain of events, transducing light into neural electrical signals.
The discs in the outer segments of the photoreceptors are regularly being formed near the cilium and phagocytized by the pigmented epithelium. Loss of this normal turnover in the outer segment may be important in several retinal diseases (e.g., retinitis pigmentosa).
Photopigments of the discs are made up of proteins, called opsins, and retinal, an aldehyde of vitamin A. The light-sensitive retinal molecule is bound to the opsin, which is a member of the G-protein-coupled membrane receptor family (see Chapter 1). When light is absorbed by a rod or cone, the retinal is transformed in a way that activates the opsin, producing a change in the concentration of an intracellular second messenger, which leads to a change in the membrane potential of the photoreceptor cell. The configurational change in retinal is the only light-sensitive event in vision.Unlike most sensory receptor cell membranes that depolarize with stimulation, photoreceptors hyperpolarize when struck by light. In rods the visual photopigment is called rhodopsin. In the dark, many gated sodium ion (Nai) channels remain open, allowing leakage of Na+ ions into the rod, which keeps the membrane in a depolarized state. When photons of light strike rhodopsin, the resulting change in second-messenger concentration produces a closing of many of the gated Nai channels. The result is a hyperpolarization of the receptor cell membrane and a decrease in transmitter released at the synapse with the bipolar cell. Photoreception in cones works in a similar manner except that the photopigment opsin is different from that of rhodopsin. Depending on the species, different populations of cones may have different opsins. Because different opsins differ in (heir ability to absorb particular wavelengths of light, the number of different cone populations in a species is related to the ability of that species to discriminate different colors.
Differences in the functional properties of rods and cones, differences in their retinal distribution, and differences in the synaptic organization of other retinal neurons to which they pass their information result in a “rod retinal system” and a “cone retinal system” that have different functional attributes. Because individual rods are more sensitive to light than individual cones, and because several rods feed their synaptic input to a single bipolar cell (convergence), the rod system is the night-vision system concerned with detecting the presence of light.
Because of the convergence of many rods on a bipolar cell, however, the rod system is not good for discerning the detail of a visual image (poor visual acuity). Also, since the rods are more highly concentrated in the peripheral retina than the cones, this region is better at detecting the presence of light than discerning its details. Finally, because all rods contain the same visual photopigment, the rod system cannot discriminate different colors.As noted, a cone is less sensitive to light than a rod. Also, one or only a few cones feed their synaptic input to a single bipolar cell. Therefore the cone system needs higher levels of illumination to function compared with the rod system. In fact, the cone system actually represents the day-vision system because the rod system does not function well in high levels of illumination. However, due to a lack of convergence onto bipolar cells, the cone system is good at discerning image details. Because the cones have a particularly high density in the region of the fovea (the central retina), the fovea is the part of the retina with the highest visual acuity. This makes sense considering the fovea is where light falls when an animal looks directly at an object of interest. Again, different populations of cones can contain different photopigments, each differentially sensitive to a range of light frequencies. Thus the cone system is also the color-vision system. Generally, the more cone photopigments present in the retina, each within a different population of cones, the better is the ability of the species to discriminate different wavelengths of light, or to “see” color. In primates there are three different populations of cones, each with a different photopigment. According to the Yoimg-Helmholtz theory, the brain assigns color to a given wavelength of light by comparing the relative activation of the different cone populations. Humans who lack one or more of these cone types» as a result of faulty genetic transmission, cannot see colors within particular ranges of the visible spectrum of light.
The most common types of such “color blindness” are linked to the X chromosome.The extent to which various veterinary species perceive color is still controversial. It is believed that mammalian ancestors may have had four different types of cones, whereas early nocturnal mammals may have had only two types of cones, having traded some of their color discrimination ability for rod light sensitivity. Most current mammals, including dogs, have only two types of cones. As noted, primates have three types of cones. It is thought that their successful exploitation of color-rich arboreal environments may have induced selection pressure for additional color discrimination ability, above that imparted by two populations of cones. Modern birds appear to have four populations of cones, one of which is sensitive to light in the ultraviolet (UV) range of the spectrum. Behavioral experiments that show their ability to discriminate UV light suggest that birds have a richer color perception than primates. Lizards, turtles, and some fish also possess UV-sensitive cones. Only primates are known to have the color vision with which humans are familiar.