PHYSIOLOGIC RESPONSES TO HEAT
1. How can the diversion of blood to the skin result in loss of body heat? How can heat loss by this means be regulated?
2. What is the stimulus for allowing heat to be lost via the skin?
3.
Where are thermosensitive cells located in the brain?4. Are there any reflexes associated with heat gain or heat loss?
5. What percent of the heat produced in the body is normally lost by insensible means?
6. What type of sweat glands predominate in animals?
7. What is the principal function of the apocrine sweat glands?
8. Is sweating an important mechanism for heat loss among domestic animals? Which one of the domestic animals represents the greatest use of this means? The least?
9. What function is accomplished by panting? What is panting? How is hyperventilation prevented while panting? Is panting only observed in the dog?
0. Which domestic animals are most able to withstand extremes of heat?
11. What are factors associated with the pig’s intolerance to heat?
2. How does the cat increase evaporative heat loss?
3. What is an approximate body temperature of birds? Why is pulmonary ventilation more likely to cool the body of birds than that of mammals?
Heat is produced constantly in the body as a result of metabolism. If there were not provisions for losing heat, the temperature of the body would increase to intolerable levels. Two principal means for losing heat are: (1) radiation, conduction, and convection and (2) evaporation of water from the skin and respiratory passageways. A third way considers the excretion of feces and urine that leave the animal at body temperature. Heat lost by excretion of feces and urine is small and is considered negligible. Under ordinary conditions, about 75% of the heat lost from the body is dissipated by radiation, conduction, and convection and is controlled mostly by vasomotor activity.
Circulatory Adjustments
Inasmuch as circulating blood is a distributor of body heat, heat can be lost from the blood if blood is brought to the skin surface and exposed to a gradient for loss to the environment.
A schematic section of the skin of,the dog (Figure 13-2) illustrates the extensive network of blood vessels to the skin. The amount of blood circulating to the skin is controlled by sympathetic vasoconstrictor fibers to the blood vessels. An increase in tone results in constriction of blood vessels and diversion of blood from the surface, thereby conserving heat. A decrease in tone lets more blood go to the surface. A stimulus for a decrease in tone so that more heat can be lost from the body is the temperature of the blood circulated to the brain. Thermosensitive cells in the rostral hypothalamus respond to warming by activating physiologic and behavioral heat loss mechanisms (Figure 13-3). Similarly, cooling of the same region stimulates other thermosensitive cells to evoke thermoregulatory responses for heat gain. Reflexes to inhibit vasoconstrictor tone also arise from thermoreceptors in the skin and other parts of the body.
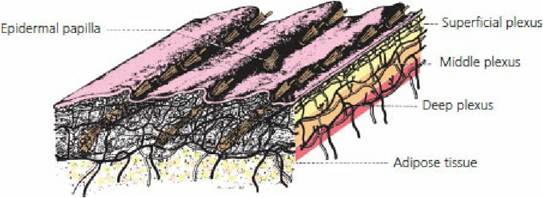
■ FIGURE 13-2 Schematic section of dog skin showing the extensive network of blood vessels and the location of insulating adipose tissue. (From Evans HE. Miller’s Anatomy of the Dog. 3rd edn. Philadelphia, PA: WB Saunders Company, 1993.)
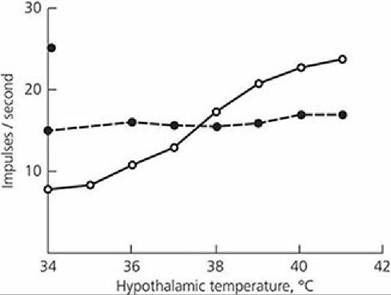
■ FIGURE 13-3 Response of warmth-sensitive neurons (solid line) in the rostral hypothalamus of the cat to increasing hypothalamic temperature. Neurons insensitive to warmth (dashed line) do not increase their activity. (From Nakayama T, Hammel HT, Hardy JD, Eisenman JS. Thermal stimulation of electrical activity of single units of the preoptic region. Am J Physiol. 1963; 204: 1122.)
Evaporative Heat Loss
Evaporation of water results in cooling.
Loss of water by evaporation is referred to as insensible water loss; this includes water lost from the skin surfaces and water lost in the heated exhaled air. Normally about 25% of the heat produced in an animal at rest is lost when water is lost by insensible means.Evaporative heat losses are increased by sweating and panting. The relative importance of sweating as a heat loss mechanism varies among species. Generally, the function of sweat glands as dissipaters of body heat is less effective in domestic animals than in humans.
There are two types of sweat glands: apocrine and eccrine. Eccrine sweat glands are those typically found in humans, but are sparse among domestic animals. In the dog and cat they occupy only the foot pad location. This area does not subserve thermoregulation; it provides for a moist surface and subsequent improved traction. Horses, cattle, sheep, dogs, and cats have apocrine sweat glands disseminated over the body surface (Figure 13-4). The composition, volume, stimulus for secretion, and function of apocrine sweat varies among species. In the dog, and perhaps in other species, apocrine sweat is a proteinaceous, white, odorless, milky fluid that is formed slowly and continuously. On the skin surface, it mixes with sebum from the sebaceous glands to form a protective emulsion that acts as a physical and chemical barrier. Characteristic animal odors arise from bacterial flora action on apocrine secretions. Heat loss from sweating (thermoregulatory function) is probably greatest in the horse, followed (in order) by cattle, sheep, dogs, cats, and swine.
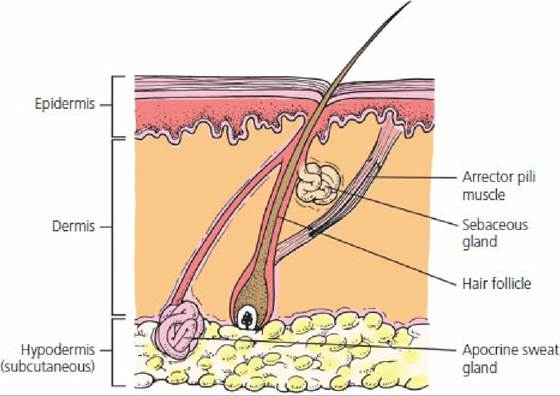
■ FIGURE 13-4 Schematic representation of apocrine and sebaceous glands and their association with a hair follicle. The secretory parts of the apocrine glands are located in the dermal and subcutaneous layers of the skin. The excretory ducts pass upward through the dermis and empty into the hair follicles above the ducts of the sebaceous glands.
The panting mechanism is effective in dissipating the heat load because greater amounts of air are made to go over moist surfaces (see Chapter 12). Panting is most effective in the dog, but it is also observed in the other domestic animals. Essentially, panting is an increase in dead-space ventilation without change in respiratory alveolar ventilation. A decreased tidal volume is associated with the increased respiratory frequency of panting; in this way, hyperventilation of the alveoli is prevented.
In cattle, panting is accompanied by increased salivation, and the salivary secretion promotes cooling by evaporation. Salivary secretion loss by evaporation and drooling (physical loss to the exterior of the body) can result in metabolic acidosis because of loss of bicarbonate and phosphate buffers contained in ruminant saliva.
Increases in sweating and panting are brought about by increased blood temperature, subsequent adjustments by the hypothalamus, and reflexes produced by local heating of the skin.
Responses to Extremes of Heat
Different animal species differ in their ability to withstand heat. The humidity of the air becomes a factor - as humidity increases, evaporation from insensible losses is reduced and less cooling occurs. Of all domestic animals, cattle and sheep seem to be the most able to withstand extremes of heat. Open-mouth panting and sweating occur as the temperature increases, and these animals can withstand temperatures as high as 43 °C (109 °F) with humidity above 65%.
The pig cannot tolerate a temperature above 35 °C (95 °F) with humidity above 65%. The intolerance of pigs to heat is recognized by transporters of livestock. During periods of heat, the transport of pigs is usually delayed until night, and they often are hosed with water. Pigs do not sweat copiously and their small mouth makes them ineffective at panting. In addition, they often have substantial subcutaneous fat.
When the relative humidity is above 65%, the cat cannot withstand prolonged exposure to an environmental temperature of 40 °C (104 °F) or higher. In addition to panting, the cat can increase evaporative losses by spreading saliva over its hair coat. Because the dog is effective at panting, it can withstand extreme environmental temperatures better than the cat, but it is in danger of collapse when its rectal temperature reaches 41 °C (106 °F).
In birds, the air sacs are extensions of the lungs, which extend into the body cavities. The body temperature of birds is about 41 °C (106 °F). Pulmonary ventilation is more likely to cool the body of birds than that of mammals because of the larger gradient and because of the closeness of the air to the body organs (via air sacs, see Chapter 10). It seems that prolonged exposure of a hen to an air temperature of,38 °C (100 °F) is unsafe if the relative humidity is above 75%. A cloacal temperature of 45 °C (113 °F) is the upper limit of safety in the chicken.
■