Physiology of the Small Intestine, Exocrine Pancreas, and Liver
The small intestine is the primary site of chemical digestion and absorption of nutrients. The exocrine secretions of the pancreas contain most of the enzymes for chemical digestion in the lumen of the small intestine, but the epithelial cells that line the small intestine (entero- cytes) also have in their cell membranes enzymes that participate in the final steps of chemical digestion.
The primary digestive function of the liver is to provide bile salts, which facilitate the enzymatic digestion of lipids. The liver is not a source of digestive enzymes.Small Intestine Secretions and Motility
Intestinal juice is derived from intestinal glands in the wall of the small intestine. These include crypts or crypts of Lieberkuhn, scattered throughout the entire small intestine (Fig. 21-4),
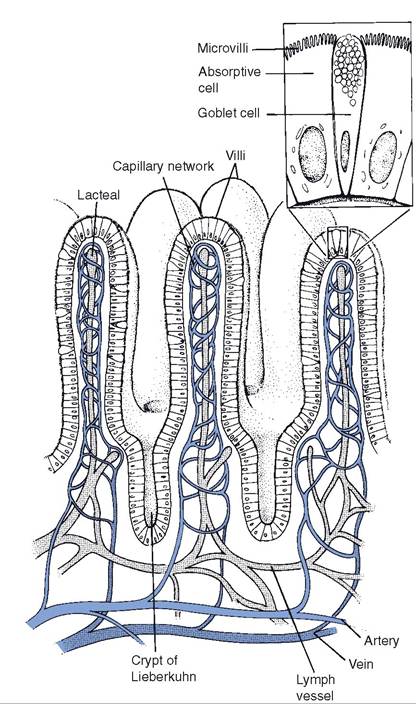
Figure 21-4. Intestinal villi and crypts. (Reprinted with permission of Wolters Kluwer from Bullock B.L. Pathophysiology. 4th ed. Philadelphia: Lippincott, 1996.)
and duodenal glands, which contribute mucus and are found only in the duodenum. The intestinal juice contains salts and water derived from blood capillaries in the wall of the intestine. The function of the secreted salts is unclear, but the water dilutes the chyme, which is usually hypertonic (higher osmolality than normal plasma). Food in the intestine stimulates secretion by these intestinal crypt glands.
The two primary types of movement by the small intestine are segmentation and peristalsis. segmentation movements, which occur when food is in the small intestine, are characterized by alternating local areas of contraction and relaxation (Fig. 21-5). These movements mix the digesta with intestinal juice and digestive enzymes and increase the contact between digesta and the epithelial surface of the small intestine.
The increased contact provides more exposure to enzymes associated with epithelial cells and to the absorptive surface of the epithelial cells. strong peristaltic contractions of the small intestine in fasting animals or several hours after a meal propel ingesta down the tract, presumably to clean the small intestine of undigested foodstuffs before the next meal.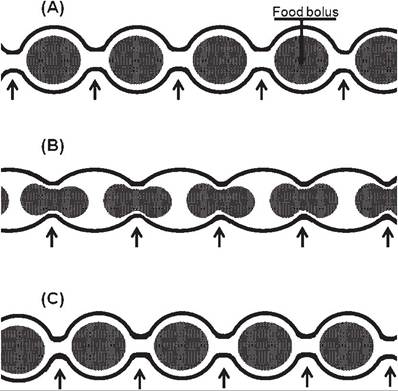
Figure 21-5. Segmentation pattern of gastrointestinal motility. The same segment of intestine is viewed at three different sequential times (A, B, and C). Regions of contractions at each time are identified by arrows.
Exocrine Pancreas
Pancreatic exocrine secretions primarily consist of a variety of digestive enzymes and sodium bicarbonate. Pancreatic acinar cells (Fig. 21-6) secrete the enzymes, and cells that line ducts in the pancreas secrete the sodium bicarbonate. These ducts empty into one or two pancreatic ducts, which empty into the duodenum.
The sodium bicarbonate raises to an acceptable pH the chyme entering from the stomach. The small-intestinal epithelium is not protected from an acidic solution by a thick layer of mucus, as is the stomach. The higher pH is also better for the action of the pancreatic digestive enzymes. The major stimulus for bicarbonate secretion is the hormone secretin from the small intestinal mucosa. secretin secretion
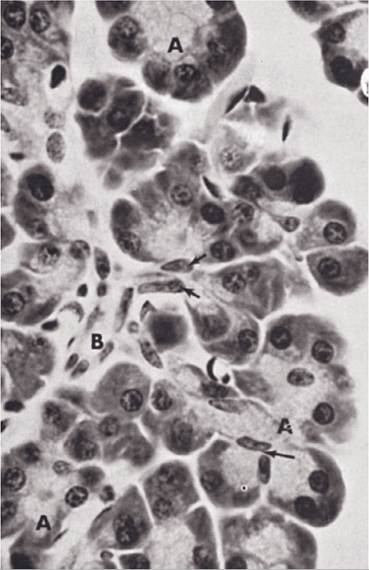
Figure 21-6. Pancreas. A, Pancreatic acini; B, intercalated duct. (Reprinted with permission of Wiley- Blackwell from Dellmann H.D. and Eurell J. Textbook of Veterinary Histology. 5th ed. Philadelphia: Lippincott Williams & Wilkins, 1998.)
increases in response to the acid chyme entering from the stomach.
Pancreatic proteolytic enzymes include trypsin and chymotrypsin. Similar to pepsin in the stomach, these are secreted as inactive precursors, trypsinogen and chymotrypsinogen.
Trypsinogen is activated by an enzyme, enterokinase, a component of the luminal cell membranes of small intestinal cells (entero- cytes). Trypsin can activate chymotrypsinogen and more trypsinogen. The ultimate end products of protein digestion are amino acids, but the pancreatic proteolytic enzymes may stop digestion when the peptides reach a length of two or more amino acids. if this occurs, peptidases associated with enterocyte cell membranes can complete hydrolysis of the peptides to individual amino acids for absorption.
Unlike the proteolytic enzymes, pancreatic amylase and lipase are in the active forms when secreted from the pancreas. Amylase digests starches to oligosaccharides (a carbohydrate composed of a small number of monosaccharides, usually two to four). The enzymes maltase and sucrase, components of entero- cyte cell membranes, further digest the oligosaccharides to monosaccharides. Lactase, to digest lactose (milk sugar), is present in entero- cytes of young mammals but not in all adults. Lipase hydrolyzes triglycerides into fatty acids and glycerol. This action is most effective after the fats have been emulsified by bile (discussed later).
Control of pancreatic exocrine secretion depends on stimulation by vagal autonomic nerves that innervate the pancreas and on three intestinal hormones, cholecystokinin, secretin, and gastrin. seeing or smelling food stimulates vagal stimulation, and food in the stomach prompts release of gastrin.
The greatest amount of pancreatic exocrine secretion occurs when the acid chyme and food components in the duodenum stimulate the release of cholecystokinin and secretin from cells in the duodenal mucosa (intestinal phase of control). These two duodenal hormones also feed back to the stomach to decrease secretions and slow down the activity and emptying of the stomach until the duodenal chyme has been degraded by the enzymes and adjusted in pH by the pancreatic bicarbonate.
Liver Digestive Function and Secretion of Bile
Liver cells (hepatocytes) are responsible for bile formation.
Bile is a greenish-yellow salt solution consisting primarily of bile salts, cholesterol, phospholipids (lecithins), and bile pigments (bilirubin). Hepatocytes synthesize the bile salts (primarily sodium salts of glycocholic and taurocholic acids) from cholesterol. These salts assist in digestion and absorption of lipids (triglycerides), and the production and secretion of these salts is the most important digestive function of the liver.in an aqueous solution, such as the duodenal chyme, lipids tend to clump together and form large droplets (recall the appearance of an oil and vinegar salad dressing after shaking the bottle). such large lipid droplets present a small surface area for the action of the pancreatic lipases. Bile acids act as emulsifiers to reduce droplet size and make the lipids more accessible to the lipases. Lipases can function without bile salts, but lipid digestion is inefficient without them. Micelle is the term for the small droplets formed in the intestinal chyme that contain lipids, bile salts, and products of lipid digestion.
in all farm animals except the horse, bile is stored in the gallbladder. since the horse has no gallbladder, the bile passes directly from the liver to the duodenum by way of the bile duct and its tributaries at a fairly continuous rate. The gallbladder stores bile for intermittent discharge into the duodenum and concentrates the bile by reabsorbing water from the stored bile. Cholecystokinin stimulates gallbladder contraction and the release of stored bile. since food entering the duodenum stimulates the release of cholecystokinin, this coordinates the release of bile with the presence of food.
Most of the bile salts released from the liver remain mixed with the digesta as it passes into the terminal part of the small intestine (ileum). Here, enterocytes reabsorb bile salts, which enter the blood. The reabsorbed bile salts are transported to the liver via the hepatic portal vein, and here hepatocytes take up the bile salts from the portal blood.
These bile salts can then be secreted by the hepatocytes into bile for reuse. An increase in bile salts in portal blood, such as during the digestion of a meal, is the primary stimulus for bile salt secretion by hepatocytes. The recycling of bile salts between the digestive tract and the liver is enterohepatic circulation.The liver is capable of synthesizing cholesterol, and the liver makes much of the cholesterol in bile. The liver can also eliminate excessive dietary cholesterol via the bile. Cholesterol is insoluble in water, but the bile salts and lecithin normally change it to a soluble form so that it can exist in the bile. However, sometimes cholesterol precipitates from the bile in the gallbladder or bile ducts, forming gallstones.
Nutrient Absorption in the
Small Intestine
The small intestine is the major site of nutrient absorption. Most of the products of carbohydrate, protein, and lipid digestion are absorbed as the digesta pass through the small intestine. The small intestine is also the primary site of absorption for vitamins, minerals, and water.
The epithelium lining the small intestine has structural features that increase the surface area for nutrient absorption. The mucosa is covered with villi (fingerlike projections) that extend into the lumen, and the individual enterocytes have microvilli on their cell membrane facing the lumen (Fig. 21-4).
individual amino acids and monosaccharides (simple sugars) are the simplest products of protein and carbohydrate digestion, respectively. The cellular mechanisms of absorption of amino acids and monosaccharides (primarily glucose) are similar in that the transport across the cell membrane on the luminal surface involves sodium-linked co-transporters (see Chapter 2). The co-transporters all bind
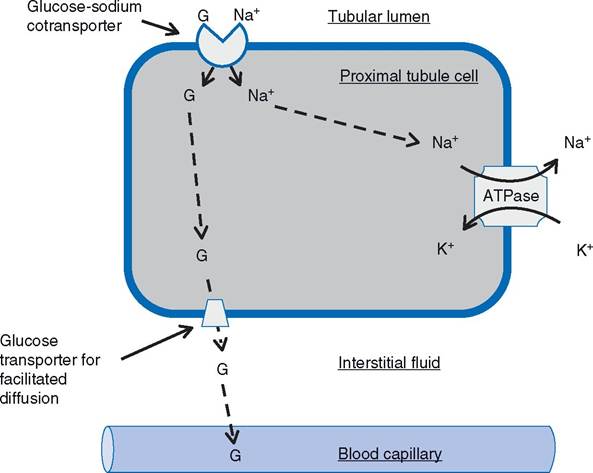
Figure 21-7. Sodium-Iinked co-transport of glucose. Similar mechanism found in proximal tubule cell (as labeled) and epithelial cell of small intestine.
Co-transporter for sodium and glucose (G) is found on the portion of the cell membrane that lines the lumen of the tubule (or gut), and the Na-K-ATPase is found on the base and lateral sides of the cell.sodium, but different co-transporters are used by glucose and amino acids. At least five cotransporters have been found to transport various amino acids. These are all characterized as secondarily active transport, for they depend on the gradient for sodium between the intracellular fluid of the enterocytes and the fluid in the lumen of the small intestine (see Chapter 2). Figure 21-7 summarizes the absorptive process for glucose.
Micelles formed in the lumen of the small intestine contain bile salts, triglycerides, cholesterol, and the products of lipase action on triglycerides (fatty acids and monoglycerides). intestinal movements (segmentation) bring the micelles into contact with the microvilli of the enterocytes. The lipid-soluble products of lipid digestion and cholesterol can then diffuse from the micelle into the enterocytes (Fig. 21-8). Some long-chain fatty acids are also transported from micelles into the cell by specific sodium- linked co-transporter proteins in the cell membrane.
Within the enterocytes, absorbed monoglycerides and fatty acids are used to resynthesize triglycerides. The enterocyte packages the triglycerides and absorbed cholesterol together with intracellular proteins into a particle known as a chylomicron. The enterocytes secrete the chylomicrons into the interstitial fluid, where they are absorbed into lymphatics (Fig. 21-8). The smallest of the lymphatic vessels that absorb the chylomicrons are Iacteals (Fig. 214). The lymphatic drainage from the intestinal tract is ultimately added to the blood, and it is via this pathway that chylomicrons, containing absorbed lipids, reach the blood.
Sodium, potassium, phosphate, calcium, chloride, and other electrolytes are primarily absorbed in the small intestine by both active and passive mechanisms. With the exception of iron and calcium, the absorption of these minerals is not regulated, so that most of what is consumed is absorbed. iron absorption is reduced at the level of the enterocyte if body iron content is sufficient. This reduction is
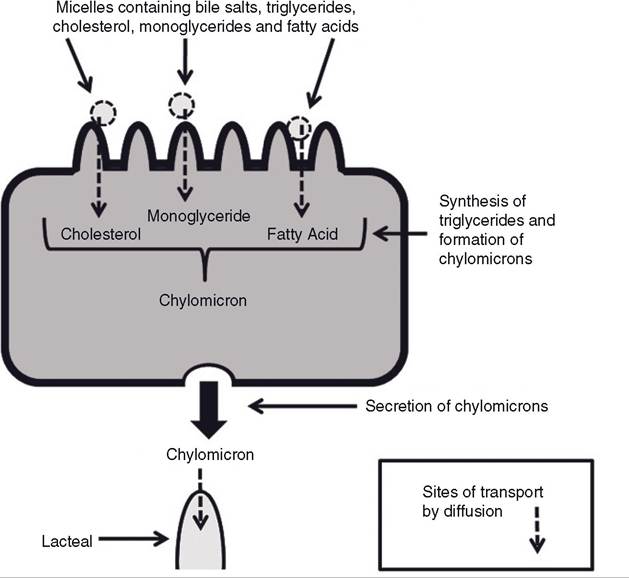
Figure 21-8. Absorption of lipids and cholesterol by intestinal cell and subsequent formation and secretion of chylomicrons.
primarily accomplished by an increase in an intracellular protein that binds iron in the enterocytes. The iron-containing enterocyte is lost from the body after it sloughs from the epithelial lining. The form of vitamin D produced by the kidneys (calcitriol) increases calcium absorption by increasing calcium transport proteins in enterocytes. Calcitriol formation by the kidneys is increased when blood calcium is low. To be absorbed, minerals such as calcium and phosphate must be in their ionized state. if the ratio of cations to anions is too high or too low, absorption can be reduced. For example, if the dietary content of the phosphate (an anion) is too high relative to calcium (a cation), the excess phosphate binds the available calcium to form calcium phosphate, and calcium absorption is impaired.
The small intestine of some neonatal (newborn) animals can absorb macromolecules, including intact protein molecules from the colostrum. Colostrum, the first milk of the horse, pig, and ruminants, contains the γ- globulins needed to produce passive immunity in the newborns of these animals. This receptive period lasts approximately 1 day in the horse and pig and up to 3 days in the ruminant.