PRENATAL DEVELOPMENTAL CHANGES AT THE CELLULAR LEVEL
Cleavage denotes a sequence of swift mitotic divisions taking place in the early embryo following fertilization. The process of cleavage in the early stages of embryonic development involves a series of mitotic divisions of the zygote, leading to the formation of a multicellular embryo.
The onset of the first cleavage involves the emergence of a cleavage furrow, manifesting as a ring encircling the zygote. This furrow aligns parallel to the polar axis (extending from the animal pole to the vegetal pole) and perpendicular to the longitudinal axis of the mitotic spindle. Comprising actin and myosin proteins, the contractile ring responsible for the cleavage furrow formation resides in the cortical region of the zygote. Myosin controls the actin filaments, enabling the contraction necessary for the furrow to deepen and eventually separate the zygote into two blastomeres. During the second cleavage, alignment parallel to the polar axis, akin to the first cleavage, is observed. As the second cleavage concludes, the embryo comprises four blastomeres arranged from the animal pole to the vegetal pole. Cleavage continues as a fractionating process, with subsequent divisions further increasing the number of cells. In the developmental sequence, the third cleavage conventionally proceeds perpendicular to the preceding two, thereby culminating in the formation of an eight-cell stage. Continued divisions lead to the formation of a 16-32 cell stage, culminating in the creation of a compact mass of cells referred to as the morula (Abbott and Ducibella, 2001). During the latter phase of the 8-cell stage cleavage, a phenomenon known as cell compaction ensues, facilitating the establishment of cell junctional complexes and imparting polarization to the cell. Consequently, the cell assumes a distinct structural orientation, with a defined contact area adjacent to neighboring cells, termed the basolateral domain, and an unencumbered outer surface known as the apical domain. Notably, the orchestration of compaction is intricately regulated by extracellular calcium ions (Ca2+), acting through epithelial cadherin (E-cad) (Dietmar et al., 1987). The apical domain accumulates apical polarity proteins such as the Par3-Par6-aPKC complex, along with cytoskeletal components, while the basolateral domain harbors proteins like Par1 and Lgl. Apical accumulation of actomycin which is facilitated by PKC activation along with proteins Par3-Par6-aPKC complex establishes polarization of the embryo (Lin et al., 2000; Vinot et al., 2005; Suzuki and Ohno, 2006). Polarization is a critical process that dictates the future differentiation and organization of the embryo’s cells. Post-polarization, blastomeres can undergo symmetric and two types of asymmetric divisions. Asymmetric division produces 1. Two polarized daughters 2. Following cleavage, one daughter cell becomes polarized while the other remains non-polarized. Non-polarized cells, situated within the blastomere’s interior, undergo differentiation into the Inner Cell Mass (ICM). The ICM possesses pluripotency, giving rise to the embryo proper and certain extraembryonic tissues. Polarized cells, positioned on the blastomere’s periphery, differentiate into the Trophectoderm (TE), which constitutes the outer layer of the blastocyst. The TE contributes to placental formation and the development of supportive tissues.The blastomere maintains a balance between ICM and TE cells by regulating the types of divisions. This balance is essential for proper embryonic development, ensuring that there are enough cells to form both the embryo and its supporting structures (Mu Jian, 2022).
By the 32-cell stage, the outer cells of the embryo become fully committed to the trophectoderm (TE) lineage. These TE cells play a vital role in forming the placenta and other supportive tissues crucial for fetal development. Na+/ K+ pumps situated on the basolateral (inner) domain of TE cells actively transport sodium ions (Na+) out of the cells and potassium ions (K+) into the cells.
Additionally, Na+∕H+ cotransporters located on the apical (outer) domain of TE cells facilitate the movement of Na+ ions into the cells and protons (H+) out of the cells. The activity of these pumps and co-transporters heightens the osmotic pressure within the intercellular spaces. This higher osmotic pressure draws water into these spaces, leading to fluid accumulation. The pressurized fluid causes the cell-cell contacts within the TE to fracture, creating hundreds of micrometer-sized lumens (small fluid-filled cavities) (Dumortier et al., 2019). The intercellular spaces merge; giving rise to a singular, larger fluid-filled cavity termed the blastocyst cavity or blasto- coel. As fluid accumulates within the blastocoel, the inner cell mass (ICM) remains attached to a specific region of the trophoblast. Subsequent to compaction and cavitation, the initial differentiation of blastomeres yields two distinct cell types: the trophectoderm and inner cell mass. In pigs, the cavitation process initiates at the 16-cell stage of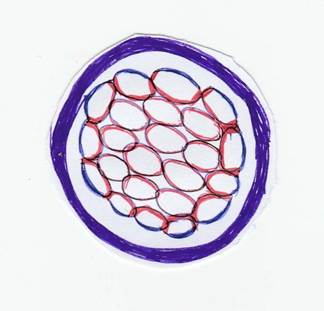
FIGURE 3.1 Events of early embryonic development
the morula. However, in humans, cattle, and sheep, cavitation commences later, at the 64-cell stage of the morula (McGeady et al., 2017) (Figure 3.1).
During early embryonic development, the first major cell fate decision occurs with the formation of the blastocyst. In cattle and sheep, blastocyst formation typically occurs around the 7th day post-fertilization. In pigs, this process usually occurs around the 5th day post-fertilization. In humans, it generally takes place by the 5th or 6th day post fertilization (Yoshinaga, 2018). After blastocyst formation, the trophectoderm contributes to placental formation, while the inner cell mass (ICM) generates the embryo proper and certain extraembryonic tissues. Immediately post-blastocyst formation, it expands and hatches from the zona pellucida. During this hatching process, cells within the ICM face a second fate decision.
This decision involves the differentiation of the ICM into two distinct cell lineages: the Epiblast (Epi), which is pluripotent and gives rise to the embryo itself, including all body tissues and organs, and the primitive endoderm, which contributes to extraembry- onic tissues, particularly the yolk sac. The yolk sac provides vital support and nutrients to the developing embryo. Specific molecular markers and signaling pathways guide the differentiation of these two cell populations within the ICM (Figure 3.2). This second fate decision is critical for proper embryonic development and the establishment of foundational structures necessary for subsequent growth and differentiation (Mu Jian, 2022).Growth and expansion of the blastocyst during the preimplantation period is species-specific. The epiblast, along
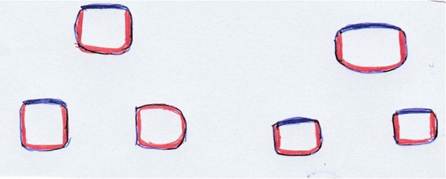
FIGURE 3.2 Cross section of early embryo showcasing polarization polarization
with the underlying primitive endoderm, forms the embryonic disc and is reshaped. In horses, dogs, cats, and rabbits, the blastocyst appears round to oval, whereas in cattle, sheep, and pigs, it expands like a filament or thread. At 9 days gestation, the porcine blastocyst appears to be a sphere of 10 mm in diameter, which lengthens up to 100 centimeters by the 13 th days of gestation. The elongation of the conceptus marks a crucial milestone in early pregnancy across these species, commencing on days 11-12 in pigs, days 12-13 in sheep, and days 13-14 in cattle. This process coincides with pregnancy recognition and implantation (Bazer et al., 2011). Interferon-Tau (IFN-τ), a pivotal signaling molecule produced by the bovine embryo, communicates with the maternal endometrium to inhibit luteolysis and sustain pregnancy. Following implantation, the developing embryo exits the uterine lumen and adheres to the uterine mucosa for the duration of gestation. This timing is crucial for synchronizing the development of the blastocyst with the receptive state of the endometrium, guaranteeing successful attachment and subsequent embryo development.
Implantation in sheep typically occurs around 15-16 days after fertilization. In cattle, it generally happens around 18-20 days post-fertilization, while in pigs, it occurs approximately 14-16 days after fertilization (Spencer et al., 2004; Anna Piliszek and Zofia E. Madeja, 2018).
Following blastulation, the embryonic disc is established, and the embryo is now all set for gastrulation. The blastula is reorganized into a three-layered structure comprising the ectoderm, mesoderm, and endoderm. Each of these germ layers will further differentiate into specific tissues and organs, laying the groundwork for the complex anatomy and functionality of the organism. Gastrulation sets the stage for organ formation by establishing the basic body plan and positioning the germ layers. Cells receive signals during gastrulation that guide their future differentiation into specific cell types and tissues. This process plays a vital role in establishing the anterior-posterior, dorsal-ventral, and left-right axes of the embryo, essential for the correct spatial arrangement of tissues and organs. Gastrulation initiates with the emergence of a primitive streak on the surface of the blastula, indicating the future posterior end of the embryo. Cells originating from the outer layer (epiblast) migrate inward through the primitive streak. These migrating epiblast cells differentiate into three primary germ layers: ectoderm, mesoderm, and endoderm. The establishment of the notochord sets the cranial-caudal axis, which is fundamental for the spatial organization of the developing embryo. The ectoderm gives rise to the epidermis of the skin and neural tissue, while the endoderm forms the lining of the gastrointestinal and respiratory tracts. From the mesodermal layer, the urogenital, circulatory, and supportive muscular and skeletal systems emerge (McGeady et al., 2017).
The completion of gastrulation marks the transition to further stages of development, such as the first embryonic organ system: neurulation (formation of the neural tube) and organogenesis (formation of organs) (McGeady et al., 2017).
In cattle, during the first two weeks of the pre-embryonic stage, the zygote is formed, and the process of implantation starts. The embryonic period commences with tropho- ectodermal adhesion to the endometrial epithelium. By its conclusion, indicated by the initiation of fetal bone mineralization, all organ systems are structured in their elementary form. Subsequently, the embryo transitions to the fetal period, spanning from the ninth week of gestation until birth canal expulsion (Valadao et al., 2018).
In sheep prenatal development, the blastocyst implants approximately 15 days after fertilization. The embryonic period extends until around the 34th day of intrauterine life, followed by the onset of the fetal period around the 35th day after conception, which continues until birth (Green and Winter, 1945; Valasi et al., 2017).
In the prenatal development, the period of the ovum spans from Days 2 to Day 17, followed by the embryo period from Days 19 to Day 35. Subsequently, the fetal period of the fetus extends from Day 35 until birth (Phemister RD, 1974).
In the prenatal development of the horse, implantation occurs around Day 16, and a series of morphological changes, embryogenesis, and organogenesis follows the embryonic period. The fetal period extends after Days 40 to birth (Ginther, 1983a, 1983b; Enders and Liu, 1991; Sharp, 2000; Allen and Wilsher, 2009; Franciolli et al., 2011).
3.4.1 Molecular and Biochemical Events
During the Development of Animals
The cellular identity is attributed to the differential expression of genes which is regulated by various factors. These expressions may determine the fate of the cell through processes like proliferation, interactions with neighboring cells, creating a special group of cells and migration of cells to a definite locus (Vestweber et al., 1987). Cellular environment along with chemical signaling plays a pivotal role in such a process. The fate of the cells is also somewhat regulated by the memory clock ticking inside it. Genes like PURA, which codes protein like Purα, are responsible for the formation of Purkinje cell layer and synapse formation in mice. Apart from that it is important for many other aspects of cellular development. This gene is ubiquitous and binds to DNA fragments containing the -GGCGGA- site present proximal to myelin basic protein (MBP) region. Any abnormality in such protein/homologous proteins may lead to defective neuronal function (Khalili et al., 2003). This indicates the importance of certain generalized factors contributing to the development in animals. Though the genetic complexity in animals is prominent, they share a significant homology between more than half of the genes. The genes which are specific to a particular organism, though are present, contribute less towards the developmental procedure. Also, regulatory genes like Nr2f, ptch, SaII, Smad6, Sp5, Tbx2/3 are universally present and conserved across different species (Frith and Ni, 2023). Some homeo- box genes like Gsx, Msx, Pitx are also conserved which bind to a conserved -CCAAT- box. Broadly, two classes of proteins are responsible for development of animals
(i) Protein spanning the membrane and ion channels
(ii) Protein that play a regulatory role.
The former is having a major role in development. The signals to create differentiation between animals are largely controlled by the regular DNA which are mostly noncoding and constitute a major portion of the genome of the animals. More than 2500 enhancers have been identified recently by Wu et al., 2022., with the help of self-transcribing active regulatory region sequence (STARR sequence) in pigs. ZPA regulatory sequence (ZRS) is an established enhancer. It is a functional element gene which is responsible for limb specific (LMBR1) sonic hedgehog (SHH) signaling molecules. Deletion of ZRS results in reduction of limbs in mice. Further, mutation affecting ZRS gene may lead to pre-axial polydactyly and triphalangeal thumb. Different species with body plans of similar kind may also have similar sets of regulatory genes.
Exposing embryos to various ionizing radiation or by transposon mediated nonspecific mutation may help in elucidating the function of a gene. The initial masses of cells of early life may be un-committed or committed or determined as per their differential potency.
Positional value of a cell shows the position specific expression of determinants. A regulatory protein known as T-box protein (Tbx transcription factor) is actively involved in this. Tbx regulates such behavior in formation of limb and heart. The members of T- box gene family bind to the sequence -TCACACCT- of DNA. TBX4 and TBX5 forms the hind and fore limb in case of chickens. Homeobox genes like Hox gene starts the signaling involving Wnt (Wingless and Int-1) signaling pathway and fibroblast growth factor (FGF)signals in formation of limbs. TBX1 and TBX2 are members of this group of protein which control rather development of testis and formation of atrioventricular canal respectively.
Proximate /inductive interaction can also change the fate of the equivalence group of cells which happens due to any local stimuli or inducers. But the ultimate outcome is also dependent upon the interplay between inducers and other regulatory proteins. Competence refers to the capacity to react to a particular inductive signal. The inductive interaction comprises two key components. The first is the inducer, or the tissue generating the signal that alters the behavior of the target tissue. The second component is the responder, which receives the signal. In rat eye tissue formation, Pax6 serves as a competence factor. Interestingly, few inductive events between different species can bring back some lost events. For example, molar mesenchyme from mouse can induce teeth in the jaw of a bird. In contrary to inducers, morphogens have long range and graded induction effect. Graded and non-uniform distribution of morphogens across the tissue forms the basis of pattern formation or morphogenesis. Morphogens like Hedgehog (Hg) and bone mor- phogenic protein (BMP) are predominant in animals. Not only proteins but non proteinaceous vitamin-A analogue can also function as a morphogen. In some cases, inhibitors of inducers play vital role in deciding the developmental fate rather than the inducers itself (Mehelen et al., 2005). “Chordin” an inhibitor of BMP /TGFβ, navigates the cells towards formation of neurons. As discussed earlier, an internal clock is also set to govern the number of times the cell should undergo a division. Sequential induction generates a diversified population in any organs. It starts with a small population and then propagate to a nearby cell population with production of various kinds of inducers (Lang et al., 2021).
Mammalian embryo needs a very less amount food due to presence of placenta which provides nourishment to it. The fertilized egg transforms into a compact form of morula with 16 cells. These cells are arranged compactly and the tight junction between the cells zips them off and acts as a barrier from the external surroundings. Adherens junctions (AJ) play important role in cell-cell cohesion (Khalili et al., 2003). Cadherins are a group of junctional proteins that initiate this process. Cadherins are classified into E (type -I) and VE (type -II) groups as per their tissue specific expressions. Further, desmosomal cadherins can be classified into desmogleins and desmocollins. Embryo compaction begins at 8 cell stage. It starts from the cell contact points and migrates forward in such a way that the neighboring cells come in the closest contact possible. Two underlying mechanisms may play role in this (i) enhanced formation of filopodia and (ii) surface contractibility mediated by actomyosin. Calcium also plays important role in compaction as it was evident that depriving cells of calcium leads to inhibition of compaction. The polarity regulators like Par3 and Par6 gets distributed to different poles side by side to the compaction. Ezrin (cytovillin), Par6 and Par3 are normally present at the apical domain and JAM-1, Na/K ATPase, E- cadherin are located in the lateral region. Intermediate filaments are joined by desmosomes. It has some additional benefits to the adherens junction like higher tensile strength, Ca++ independent adhesive state and plasticity to stretch higher order magnitude of the length spanning. This desmosome assembles initially at 32 cell stage. During gastrulation, junctional proteins like plakoglobin and pla- kophilin acts as bridging protein for different other proteins involved in such interactions. Further, these junctions are supported by tight junctions or “Zonula Occludens(ZO)”. These tight junctions provide space for paracellular diffusion and regulate the passage of different ions and solutes. They also contribute to the polarity by compartmentalizing the plasma membrane to the apical and basolateral parts. The tight junction is made up of junctional adhesion molecules claudin and occluding. ZO (1,2,3) acts as scaffolding proteins (Canse et al., 2023). The epithelium and mesenchyme interact to generate branching structures like lungs. This is otherwise known as “branching morphogenesis”. This mostly needs modulation of “tyrosine kinases” mediated signaling. The ligands associated are well expressed in the mesenchyme and cognate receptors are expressed in the epithelium (Lang et al., 2021). Signaling facilitated by glial cell line-derived neurotrophic factor (GDNF) regulates branching morphogenesis in the kidney, while fibroblast growth factor (FGF-10) mediates this process in the lungs (Lopez et al., 2023).
3.4.2 Maternal to Zygote Transition
It is the transition point where the control passes from the mother to the zygote (Li et al., 2013). It starts with eliminating the proteins and mRNA received from mother and synthesizing own protein and RNA. Micro RNA like miR- 202-3p is responsible for elimination of these maternal proteins. The cell division increases cells number but potency of the cell decreases which renders the emergence of germ layers. Eomesodermine or T-box brain protein 2 (EOMES) are a group of transcription factors which are essential for development of eomesoderm in vertebrates. A deliberate deletion of “eomes” in mice may result in lethality due to neurodegeneration. RNA binding proteins like Bicaudal-C (Bicc1) are also involved in MZT. Bicc1 regulates the left right patterning in case of mouse and anterior posterior polarity in lower vertebrates like Xenopus. Various growth factors also play critical role in development process. Insulin like growth factor (IGF) plays a crucial role in production of embryo. These are proteinaceous factors with significant homology to insulin. Expression of IGFs is well reported in bovine endometrium, pig blastocysts and mouse blastocysts. The role of leukaemia inhibitory factor (LIF) is quite prominent in mice during implantation. TGFα and FGF interact with receptor tyrosine kinases. In mice and rats, its importance in embryo formation has been well documented (Heyner et al., 1993).It can be said that the molecular and biochemical trigger for cellular development is widely regulated through various growth factors, transcription factors and stimulants/ inducers secreted temporally and spatially.
Synchrotron X-ray imaging has revolutionized the study of biological structures and functions. Opening new avenues for research in various biological fields, from molecular biology to comparative anatomy, is the ability to provide detailed, three-dimensional images. (Westneat et al., 2008)