Respiratory System
The avian respiratory system is dramatically different from the mammalian counterpart in that the lungs are part of a unidirectional circuit that includes a series of nonrespiratory (that is, lacking gas exchange surfaces) air sacs (Fig.
30-4), and airflow is created by movements of the sternum rather than through contraction of a muscular diaphragm. The arrangement is an especially efficient way of extracting oxygen from air, in keeping with the extreme respiratory demands created by flight.The avian nasal cavity is widely open to the oral cavity via the choanal cleft, but is otherwise not functionally different from the mammalian nose. The oral end of the trachea is guarded by the larynx, which in birds comprises only cricoid and arytenoid cartilages. The opening into the larynx, the aditus laryngis, is situated in a prominence caudal to the tongue called the laryngeal mound. The avian larynx is a protector of the airway and is not the organ of phonation in birds.
The avian trachea is constructed of tracheal rings that, unlike the C-shaped rings of the mammalian trachea, are complete cartilaginous circles that are joined by annular ligaments, themselves so short that adjacent rings may overlap. The trachea bifurcates into the two principal bronchi within the body cavity. The distal end of the trachea and the first portion of the bronchi are modified into a uniquely avian organ, the syrinx. The syrinx is the avian organ of phonation. Male ducks and swans possess an additional dilation off the left side of the syrinx, believed to be a resonator.
The lungs of birds are unlobed and lie adjacent to and deeply imprinted by the thoracic vertebrae. The primary bronchus passes through the lung and at its caudal border is continuous with the abdominal air sac (Fig. 30-4). Within the lung, it gives off many small groups of secondary bronchi, which themselves give rise to hundreds of parabronchi.
Most of these are arranged in tightly parallel loops, and others form more irregular loops. These parabronchi are the site of gas exchange. The secondary bronchi are also continuous with other air sacs of the body cavity (Fig. 30-4).Air sacs are blind-ended expansions of the respiratory tree the walls of which are composed of connective tissue covered externally by serosa. They are lined by a simple squamous epithelium and are relatively poorly vascularized. In many locations, the air sacs surround and are attached to the organs within the body cavity. Some air sacs have diverticuli that extend into the medullary cavity of bones; bones thus modified are said to be pneumatic bones. Domestic fowl, in which flight is somewhat limited, have fewer pneumatic bones than species with good flight capabilities. Fractures of pneumatic bones can lead to air sac infection.
Depending on whether or not certain compartments are considered separate sacs, most domestic birds are considered to have eight or nine named air sacs. These include cervical, clavicular, cranial, and caudal thoracic and abdominal air sacs; some of these are paired.
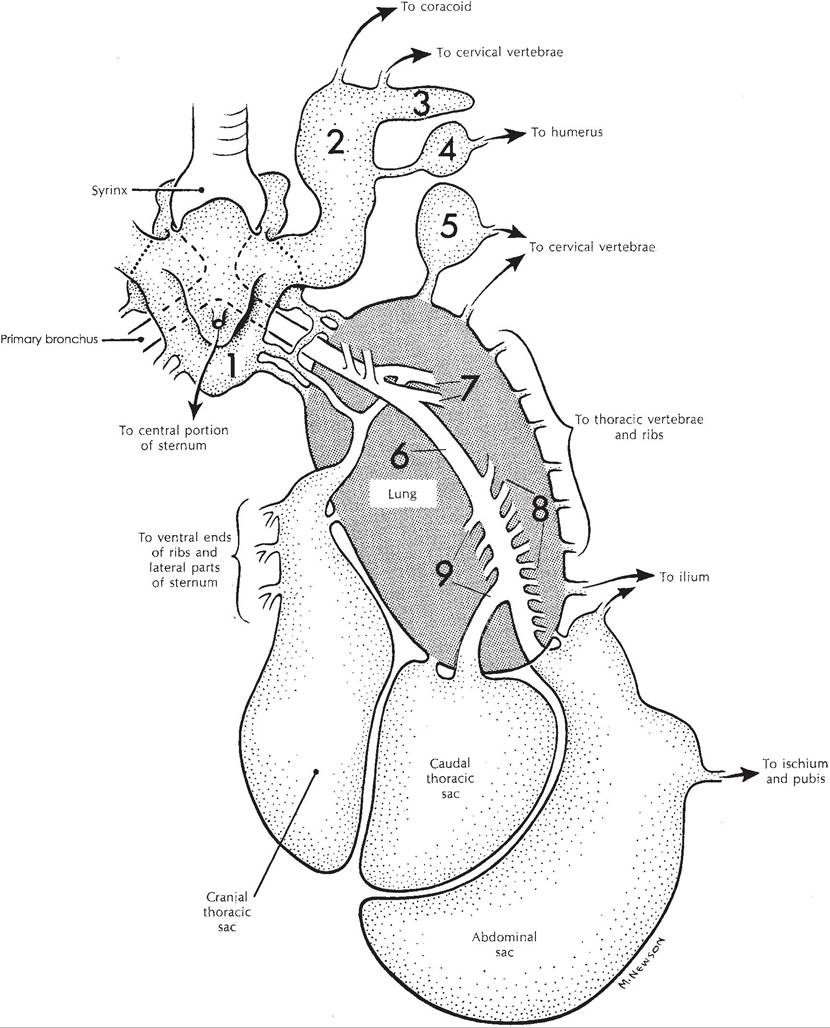
Figure 30-4. Schematic of the avian respiratory system. The trachea (unlabeled) ends at the syrinx near the top of this figure. 1, Clavicular sac; 2, lateral diverticulum of clavicular sac; 3, subscapular diverticulum; 4, axillary diverticulum; 5, cervical sac; 6, primary bronchus; 7-9, secondary bronchi. (Reprinted with permission of Wolters Kluwer from Rosskopf, W. and Woerpel, R. Diseases of Cage and Aviary Birds, 3rd ed. Philadelphia: Williams & Wilkins, 1996.)
Ventilation and Gas Exchange
Similar to domestic mammals, air movement into and out of the respiratory tract of birds requires skeletal muscle contraction. However, as noted above, avian lungs are not contained in a closed thoracic cavity and change little in size during ventilation.
Gas exchange by diffusion does occur within the avian lung, but air moves through the lungs as a result of pressure differences between the air passages within the lungs and the avian air sacs.Like domestic mammals, birds have skeletal muscles that, based on their function, are considered to be either inspiratory or expiratory muscles. Contraction of inspiratory muscles in birds results in a cranial and ventral movement of the sternum and a cranial movement of ribs of the thoracic cage. These movements enlarge the body cavity in which the air sacs are located. The enlargement results in a lowering of atmospheric pressure within both the cranial and caudal air sacs, and the resulting pressure gradient promotes the movement of outside air into the air sacs via the trachea and upper airways (Fig. 30-4). Contraction of expiratory muscles moves the sternum and ribs in the opposite direction to reduce the size of the body cavity, which increases atmospheric pressure within the air sacs and promotes an outward movement of air. unlike domestic mammals, in which expiration at rest is typically a passive event associated with relaxation of inspiratory muscles, a normal respiratory cycle in resting birds involves the appropriately timed cyclic contraction and relaxation of both inspiratory and expiratory muscles. Movements of the sternum alone are sufficient to support ventilation, and these movements can be performed manually to maintain ventilation in overly anesthetized birds. When restraining birds, care must be taken not to overly restrict the movements of the sternum, for this would significantly impair ventilation.
Air moves into both cranial and caudal air sacs during inspiration and out of both during expiration (Fig. 30-4). However, during both inspiration and expiration, air also flows through the lungs in a caudal to cranial direction via parabronchi. The unidirectional airflow through the lungs during both phases of the respiratory cycle is a unique feature of airflow patterns in the avian respiratory tract and contributes to the efficiency of gas exchange (described in a later section.) The unidirectional flow in the lungs may be maintained by local differences in airway branching patterns, airway resistances, flow velocities, and other factors, but no anatomical valves have been found.
Air capillaries are the smallest blind-ending airway of a branching system of airways extending from a parabronchus. Figure 30-5 is a simplified model of the branching system and its relationship to blood vessels for gas exchange. The connecting air passages permit gas exchange by diffusion between the lumen of a parabronchus and the surrounding air capillaries (Fig. 30-5). This exchange occurs throughout both phases of a respiratory cycle, as air flows in the same direction through the parabronchus during both inspiration and expiration as described earlier.
Air capillaries are surrounded by a meshwork of blood capillaries, a relationship similar to that for alveoli and pulmonary capillaries of the mammalian lung. In Figure 30-5, blood in the vessels entering the lungs is “venous” blood that has a relatively high partial pressure of CO2 (PCO2) and a low partial pressure of oxygen (PO2). These vessels branch so that each air capillary is perfused with venous blood with similar values of PCO2 and PO2. In this model, gas exchange can occur at all gas capillary/vas- cular capillary pairs, even though the partial pressures of O2 and CO2 are quite different in the various gas capillaries. Due to gas exchange between air and blood at each gas capillary/vas- cular capillary pair and diffusional mixing between the multiple gas capillaries and the single parabronchus, PO2 and PCO2 within the lumen of the parabronchus will decrease and increase, respectively, as air flows from beginning to end of a parabronchus. Thus, air capillaries at the end of a parabronchus will contain air with a relatively low O2 and a relatively high CO2 compared to the air in the first air capillary
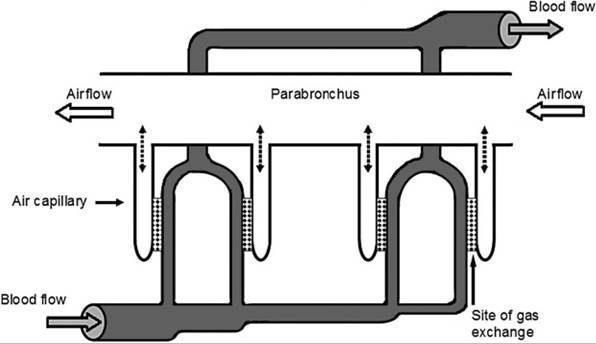
Figure 30-5. Model system illustrating direction of airflow and gas flow relative to exchange sites in avian lung.
Dotted lines between parabronchus and air capillaries represent the path for diffusion of air between these two chambers.branching from the parabronchus. However, all air capillaries are perfused by venous blood with an even lower PO2 and an even higher PCo2, so gas exchange can continue, even at the last air capillary (Fig. 30-5). Note that when blood flows past the site of gas exchange, it is flowing perpendicular to the direction of airflow within the associated parabronchus. This arrangement of blood flow perpendicular to airflow is termed a cross-current exchange model. Air exchange can also occur during both inspiration and expiration, because airflow through the parabronchi from caudal to cranial air sacs (Fig. 30-4) continues during both phases of the respiratory cycle. These factors contribute to a very efficient gas exchange, and greater percentages of oxygen in inspired air are extracted than are possible in mammalian lungs.