RUMINANT METABOLISM
1. What are the principal nonhexose sources of glucose for ruminants? Why do they need nonhexose sources? Why do they need glucose?
2. Study energy production, entry routes of the VFAs into the tricarboxylic acid cycle, ketone production, and treatment rationale for ketosis.
3. What is a summary statement for why bloat occurs?
Several indispensable uses for glucose in the body include its function as the principal source of energy for the brain. Glucose also serves as a precursor for glycerol (needed for the synthesis of fats) and as a reducing agent (in the formation of the reduced form of nicotinamide adenine dinucleotide phosphate) in the degradation of fats. In addition, muscle glycogen is formed from glucose, which serves as an anaerobic energy source during exercise. Liver glycogen present in newborn animals is derived from maternal glucose, and milk sugar (lactose) and milk fat require glucose (glycerol derived from glucose for milk fat) for their synthesis during lactation.
Gluconeogenesis
In ruminants, dietary carbohydrates (including cellulose) are fermented in the rumen to VFAs (acetic, propionic, and butyric acids) and only small amounts of glucose are absorbed. To provide glucose for the uses mentioned above, glucose must be formed (glucogenesis) from nonhexose (nonsugar) sources. In ruminants, about 85% of the glucose is formed in the liver from nonhexose sources. The principal nonhexose sources for glucogenesis in ruminants are propionate (a VFA), glycerol, lactate, and protein (via amino acids). Propionate is the only VFA that can be used for glucogenesis. It is the major source of glucose and glycogen in the ruminant (accounting for about 70%). Protein is the next most important source, accounting for about 20% under normal conditions and up to about 50% during starvation (in which propionate can be absent).
A metabolic scheme showing the pathways for glucose and glycogen formation from these four sources is shown in Figure 12-43.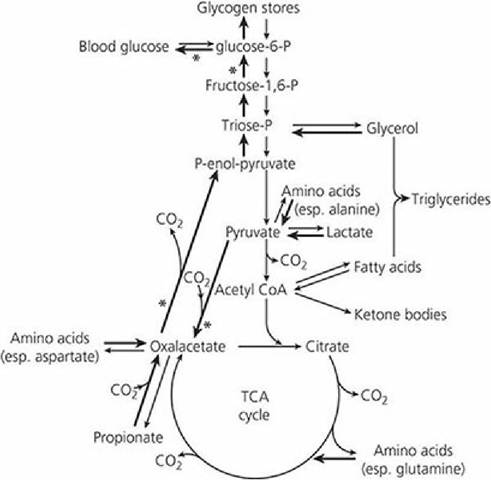
■ FIGURE 12-43 Major metabolic pathways in the ruminant liver. Because insufficient glucose is absorbed, one of the main functions of the liver is gluconeogenesis. These reactions are shown by heavy arrows; four major pacemaker reactions are indicated by asterisks. TCA, tricarboxylic acid. (From Bergman EN. Disorders of carbohydrate and fat metabolism. In: Swenson MJ, Reece WO, eds. Dukes’ Physiology of Domestic Animals. 11th edn. Ithaca, NY: Cornell University Press, 1993. Used by permission of the publisher, Cornell University Press.)
Energy Production
Two major stages are involved in the process by which energy becomes available to an animal: (1) a preliminary conversion of the proteins, carbohydrates, and fats of the diet either to acetyl coenzyme A (CoA) or to an intermediate of the citric acid cycle and (2) the subsequent oxidation of these relatively simple compounds. The second stage is represented by the citric acid cycle, also known as the Krebs cycle or tricarboxylic cycle (Figure 12-44). Propionate either can enter the tricarboxylic acid cycle as an intermediate or can be glucogenic (see Figures 12-43 and 12-44). The other VFAs (acetate and butyrate) are not glucogenic, but provide energy by entering the citric acid cycle as acetyl CoA. Acetate and butyrate (and any other substrates that enter the citric acid cycle as acetyl CoA) cannot enter the cycle unless sufficient oxaloacetate is present for condensation with acetyl CoA to become citrate. Oxaloacetate is derived from three carbon (3-C) compounds such as propionate and pyruvate or from other intermediates in the cycle. When not enough 3-C compounds are available for the formation of oxaloacetate, or if the production of acetyl CoA is excessive (e.g., in excess oxidation of fat for glucose or energy), acetyl CoA accumulates as acetoacetyl CoA, which is subsequently degraded to acetoacetate, b-hydroxybutyrate, and acetone (see Figure 12-44).
The latter three compounds are referred to as ketone bodies; accordingly, acetate and butyrate are considered to be potentially ketogenic. The condition in which excess ketone bodies are formed is referred to as ketosis.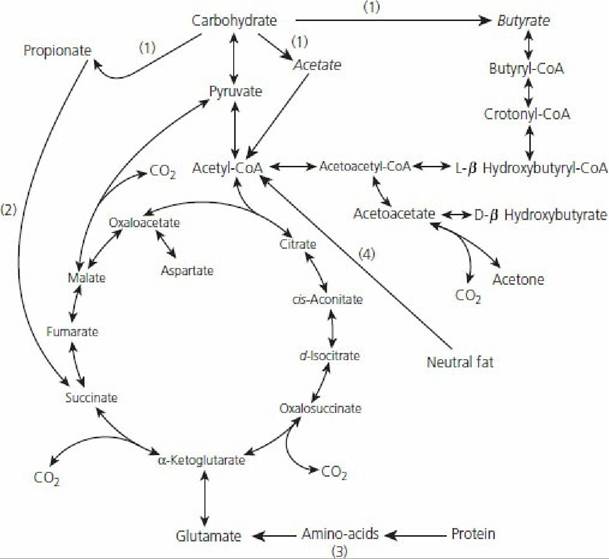
■ FIGURE 12-44 Citric acid cycle (tricarboxylic acid cycle, Krebs cycle) in relation to ruminant metabolism. Only major intermediates are shown. (1) Produced by ruminal fermentation; (2) pathway established in rumen epithelium; (3) arginine, proline, hydroxyproline, and histidine probably enter the citric acid cycle after conversion to glutamate, while aspartate gives rise to oxaloacetate by transamination; (4) the glycerol arising from the breakdown of neutral fat is metabolized by the glycolytic pathway. (From Annison EF, Lewis D. Metabolism in the Rumen. New York: John Wiley & Sons, 1959.)
Ruminant Ketosis and Bloat
In ruminants, ketosis and an associated hypoglycemia (low blood sugar level) occur most frequently in high-producing dairy cows (usually within 6 weeks after calving), in which it is called acetonemia, and in late-gestation pregnant ewes, in which it is called pregnancy toxemia. Ketosis is the end result of the animal being in a situation where the caloric demand is greater than the dietary intake. Initial efforts by the animal are to spare glucose consumption and provide alternative energy sources. This results in the mobilization of stored fat, which can be converted by the liver to acetyl-CoA for entry into the citric acid cycle (Figure 12-44). The ruminant liver has a limited capacity to convert fat into acetyl-CoA and is soon overwhelmed, at which point the mobilized fat is converted to ketone bodies and triacylglycerol. Peripheral tissues use ketone bodies as an energy source, but their ability to do so is also limited, and with continued ketone production the blood concentration will increase. Elevated levels of blood ketone levels will decrease the animal’s appetite, further contributing to the condition.
Elevated liver triglyceride (fatty liver) impairs liver function, including gluconeogenesis. In cows with clinical ketosis, there is a sudden or gradual loss of appetite, a rapid decrease in condition, and usually a marked decrease in milk flow. Although the onset is usually sudden, there may be a history of unthriftiness or a gradual loss in condition or milk production over a period of 1 to 4 weeks.The treatment of primary ketosis in cattle is directed toward increasing the blood concentration of glucose. This can be done by intravenous infusion of glucose. The feeding of sucrose (table sugar) is usually ineffective because it is quickly fermented (first to glucose and fructose and then to VFAs). Glucose precursors, such as propylene glycol, are also given as an oral drench for several days. The propylene is absorbed across the ruminal wall and used by the liver to produce glucose. Administering glucocorticoids can help elevate blood glucose levels by decreasing tissue uptake of glucose, increasing gluconeogenesis, and also suppressing milk production. Glucocorticoids should be used cautiously as they do have immunosuppressive effects and can predispose the animal to infection.
A condition of tympanism or bloating occurs when gas is unable to escape the rumen. This can result from physical obstruction of the esophagus, alteration of nerve input to the digestive system, or by the ingestion of certain feedstuffs. Two general types of feed induced bloat are recognized: (1) feedlot or grain bloat, which occurs in cattle as the result of feeding a high-concentrate diet, and (2) legume bloat, which can occur when cattle feed on lush, rapidly growing alfalfa or clover pastures. It is believed that these dietary bloats occur because the gas becomes trapped in tiny bubbles (frothy bloat) and the normal free gas bubble cannot accumulate on top of the ingesta in the dorsal sac of the rumen. The mechanoreceptors are effectively covered and the presence of gas is thus not detectable, which ordinarily would initiate the eructation reflex.
Tiny bubbles are formed because the surface tension of the rumen fluid has been increased by saponins present in legumes or in the mucopoluysaccharides some rumen amylolytic bacteria make. Surface-active agents (surfactants) have been used to lower the surface tension effectively and cause the bubbles to coalesce. As long as the gas produced in the rumen and reticulum is permitted to collect in the dorsal part of the rumen without frothing, and as long as the eructation mechanism is functioning, no problem arises from gas production, even at high rates. Problems occur when gas cannot be eliminated. Bloat does not occur because of any change in the gas composition or because of any increase in the rate of gas production, but because of a failure of the eructation mechanism. When the rumen and reticulum are distended with gas (tympanites, or bloat), pressure is applied in all directions, but becomes serious when pressure on the diaphragm prevents thoracic enlargement (needed for inspiration) and pulmonary ventilation is severely impaired.Bloat has been variously described as being caused by the highly soluble protein content of the rumen, mucin in saliva, insufficient amount of saliva, bacterial slime, the high saponin content of ingested plants, and specific eructation inhibitors. It seems that bloat might have several causes. Eating alfalfa or clover pastures (legumes) often causes bloat. However, birdsfoot trefoil, a leguminous plant, does not cause bloat.
■