Ruminants Exist in a Perpetual State of Gluconeogenesis Because of Their Unique Digestive Process
Most carbohydrate digestion in ruminants occurs in the forestomach through fermentative digestion. The result is that
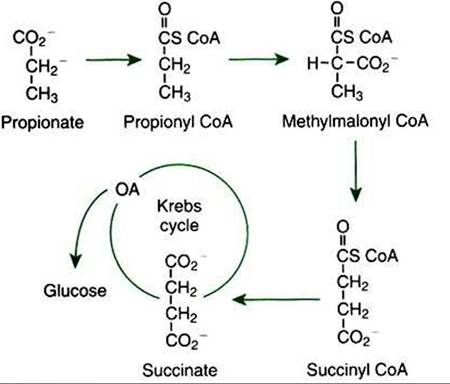
FIGURE 32-18 Gluconeogenesis from propionate involves its initial conversion to succinate.
Succinate is a four-carbon Krebs cycle intermediate that can lead to net glucose synthesis.stressed. Dairy cows are especially vulnerable in early lactation because the synthesis of lactose (milk sugar) requires glucose. In high-producing cows, nearly all the glucose they produce goes to lactose synthesis, whereas the remaining tissues function on alternative fuels. Sheep experience a similar stress on glucose synthesis in late gestation. The energy needs of the fetus and placenta can be met only by glucose (or glucosederived lactate) and amino acids. Compared with many other animals, sheep have a high ratio of fetal mass to body size; thus their fuel homeostatic mechanisms are particularly stressed by pregnancy. Failure of the glucose homeostatic mechanism frequently occurs under these circumstances, resulting in conditions known as lactational ketosis in dairy cows and pregnancy toxemia in ewes.
almost no digestible carbohydrate enters the intestine for glandular digestion and absorption as glucose. Therefore, ruminants exist in a constant state of potential glucose deficiency. To cope with this situation, ruminants have developed efficient systems of both production and conservation of glucose.
Essentially, all the glucose available to ruminants with typical diets originates from gluconeogenesis. Quantitatively, the most important glucose precursor is the volatile fatty acid (VFA) propionate. Propionate contributes to glucose synthesis after entering the Krebs cycle at the level of succinate (Figure 32-18).
Note that succinate is a four-carbon Krebs cycle intermediate that can lead to net formation of oxalo- acetate, the entry metabolite for gluconeogenesis. The other VFAs, acetate and butyrate, also enter the Krebs cycle, although they enter as acetyl CoA. As previously discussed, acetyl CoA cannot lead to the net production of Oxaloacetate or glucose. Therefore, of the ruminant’s major energy sources—acetate, propionate, and butyrate—only propionate can support glucose production. Almost all propionate absorbed from the rumen is extracted from the portal blood by the liver, never entering the systemic circulation.In addition to constant gluconeogenesis, ruminants also support their glucose needs by efficiently conserving glucose. Fatty acids are synthesized in the liver of some animals (e.g., primates, rats, dogs) but only in the adipose tissue of ruminants. Furthermore, glucose is essentially not used for fattyacid synthesis. Rather, fatty acids are synthesized from acetate, which is the most abundant energy source in ruminants. The only glucose used by adipose tissue is for the synthesis of the glycerol backbone for triglycerides. In lactating animals, fatty acids produced in the udder for milk fat are synthesized from either acetate or ketone bodies, never from glucose.
Some important metabolic diseases of ruminants occur during periods when their system of glucose homeostasis is