Sexual Receptivity Is Keyed by the Hormones Estrogen and Gonadotropin-Releasing Hormone in the Female and Testosterone in the Male
As indicated previously, the establishment of sexual behavior depends on exposure, or lack of exposure, of the hypothalamus to testosterone during the early neonatal period. In essence, testosterone (aromatized to estrogen) causes masculinization of the sexual centers in the hypothalamus; in the absence of testosterone, the hypothalamus becomes feminized.
An area within the hypothalamus, the medial preoptic area, has been identified in the rat as an area that is modified structurally by exposure to testosterone.Several principles exist regarding the effects of hormones on sexual behavior of domestic animals. First, the magnitude of change in hormone concentration that affects sexual behavior is small; in the cat, for example, an increase in estradiol-17 concentration from 10 to 20 pg∕ml. of plasma results in signs of proestrus. Second, synergism between hormones is often important for sexual receptiveness; in the dog, for example, estrogen priming followed by progesterone is important. Third, the sequence of exposure to hormones can be important; in the ewe, for example, progesterone priming is required before estrogen exposure for manifestation of estrus.
Estrogen, from the developing antral follicle, is the one hormone required for sexual receptivity in all domestic animals. Progesterone, derived from either the granulosa of the preovulatory follicle or the CL, is also important for estrus in some animals.
In sheep, estrus occurs in response to estrogen only if the animal has been exposed previously to progesterone (through the presence of a previous CL). Estrus usually begins within a short period after the end of the luteal phase (i.e., 24-36 hours) because of the presence of large antral follicles at Iuteolysis; thus the period from last exposure to progesterone and the onset of estrus is short (Figure 37-5). The requirement of progesterone for sexual receptivity means that the first follicular phase of the breeding season, which leads to ovulation in the ewe, is not accompanied by estrus.
Most adult ewes show estrus after the first luteal phase. Ewe lambs often require the exposure of two or more luteal phases before they express estrus.Of the domestic species, dogs are unusual in that sexual receptivity is keyed by progesterone, produced initially by the granulosa during the preovulatory luteinizing hormone (LH) surge and subsequently by the developing CL. Prior exposure to estrogen makes the female attractive to males but does not produce sexual receptivity; estrus requires the additional exposure to progesterone. Estrus is often maintained for up to a week in the presence of a developing luteal phase. In other domestic species, progesterone is inhibitory for estrous activity.
The importance of prior progesterone priming for estrus manifestation has been suggested for dairy cattle by the finding of a reduced incidence of estrus al the first postpartum ovulation (days 15-20). Complete progesterone withdrawal occurs in the cow immediately before delivery, and animals would not have been exposed to progesterone for 2 to 3 weeks in this situation. Sows also have a reduced incidence of estrus at the first ovulation, which usually does not occur until after weaning, usually not until at least 45 days after parturition. Other domestic species (i.e., cats, goats, horses) all show estrus with the first ovulation of the season with no apparent requirement for progesterone priming.
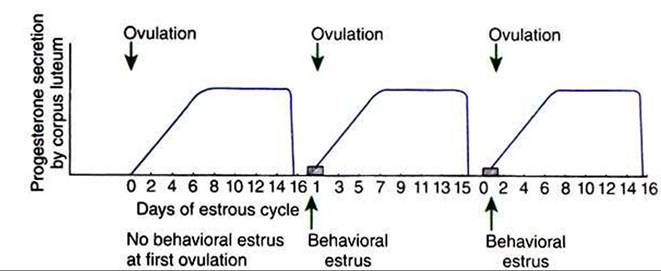
FIGURE 37-5 Estrous cycle of the ewe, showing how the first ovulation of the season is unaccompanied by estrus. Note the short interval between regression of the corpus Iuteum and the next ovulation. (From Short RV: Oestrous and menstrual cycles. In Austin CR, Short RV, editors: Reproduction in mammals, vol 3, Hormonal control of reproduction, Cambridge, UK, 1984, Cambridge University Press.)
Testosterone is important for libido in female primates. The theca layer from degenerating follicles forms an active interstitium that secretes the androgens androstenedione and testosterone.
Androgens are also essential for the maintenance of libido in males. Occasionally» castrated males, particularly horses, are able to maintain libido despite the lower concentrations of androgens (of adrenal origin) that are present after castration. These animals can sometimes be differentiated from those with retained testicles (cryptorchid animals) by testosterone analysis of plasma; however, serum testosterone levels in intact males vary by the minute. A GnRH stimulation test more accurately identifies remaining testicular tissues (2.2 μg∕kg intravenously, sampling before and 1-3 hours later). When commercially available, serum LH levels are better for differentiating bilaterally cryptorchid individuals (LH 1 ng∕mL).Both experimental and circumstantial evidence indicates that GnRH plays a role in sexual receptivity. The administration of GnRH to Ovariectomized rats produced sexual (lordotic) responses, and in prepubertal gilts, GnRH administration resulted in the occurrence of estrus within 24 hours. The circumstantial evidence is that the onset of sexual receptivity in animals is tightly coupled to the onset of the preovulatory
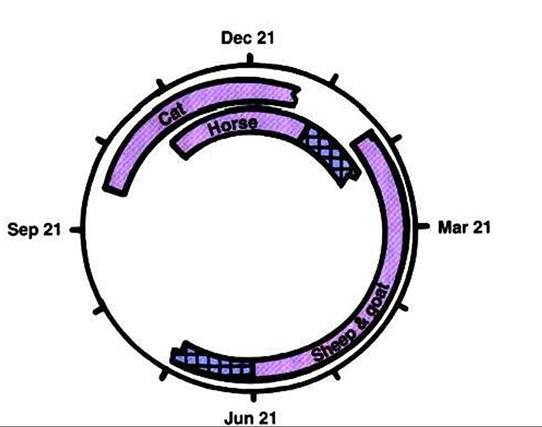
FIGURE 37-7 Diagrammatic representation of the effect of photoperiod on ovarian activity in the typical cat, horse, sheep, and goat.The bars represent periods of ovarian inactivity (anestrum).The transitional periods for the horse, sheep, and goat are shown by the hatched portions of the bars. (From Stabenfeldt GH, Edqvist LE: Female reproductive processes. In Swenson MJ, editor: Dukes' physiology of domestic animals, ed 10, Ithaca, NY, 1984, Cornell University Press.)
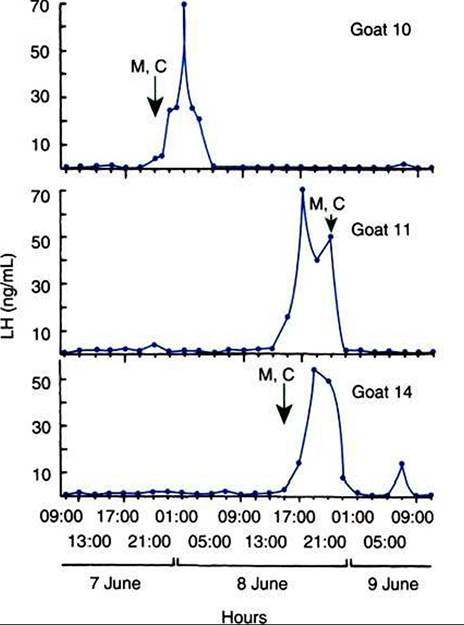
FIGURE 37-6 Plasma luteinizing hormone (LH) levels in three nannies exhibiting an ovulatory surge during the 50-hour period of intensive sampling (7-9 June). Mt Marked; C, Copulation. (From BonDurant RH, Darien BJ, Munro CJ, et al: Photoperiod induction of fertile oestrus and changes in LH and progesterone concentrations in yearling goats [Capra hircus], J Reprod Fertil 63:1, 1981.)
gonadotropin surge (Figure 37-6). Because the preovulatory gonadotropin surge is the result of an increased rate of pulsatile release of gonadotropins driven by increased GnRH synthesis and release, it is likely that this increased GnRH secretory activity affects sexual centers within the hypothalamus for the promotion of sexual receptivity. This allows the onset of the ovulatory process, triggered by the gonadotropin surge, to be tightly coupled with sexual receptivity.