Skeletal Muscle
Structure
The skeletal muscle fiber (also called voluntary striated muscle fiber) is actually a long, multinucleated cell with visible striations. Immediately beneath the outer cell membrane (sarcolemma) are numerous nuclei, reflecting the end-to-end fusion of shorter primitive muscle cells during development.
The interior of the fiber is packed with elongated protein strands (myofibrils), and filling the clefts and spaces between these strands is an extensive network of smooth endoplasmic reticulum (sarcoplasmic reticulum) and associated tubular invaginations of sarcolemma (transverse tubules, or T tubules) (Fig. 8-1).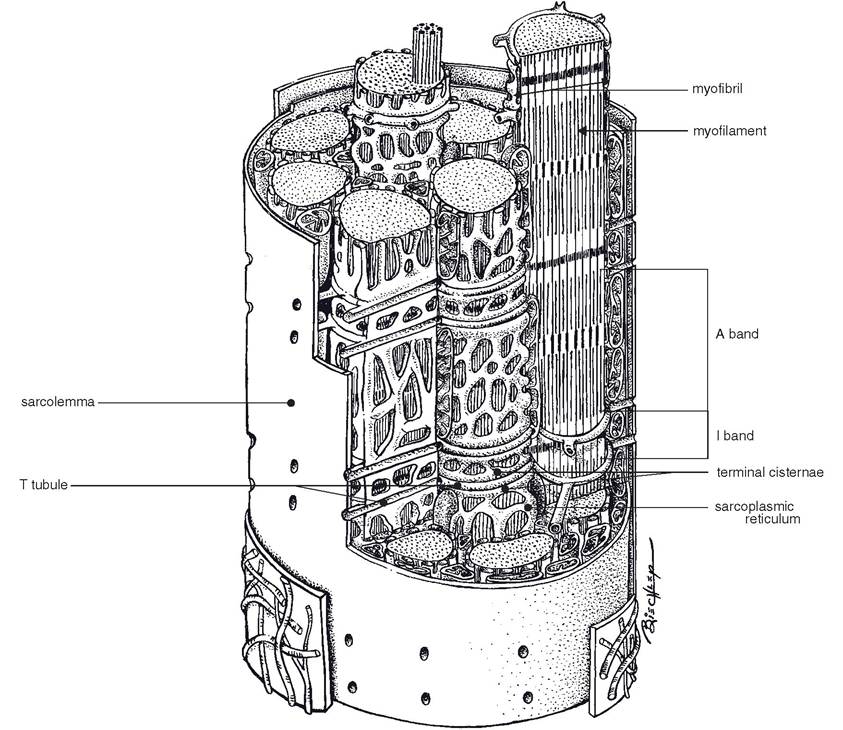
Figure 8-1. The myofibrils of a skeletal muscle cell are surrounded by sarcoplasmic reticulum. T tubules extend into the sarcoplasm from the sarcolemma to surround the myofibrils. (Reprinted with permission of Wiley-Blackwell from Dellmann D. and Eurell J. Textbook of Veterinary Histology. 5th ed. Baltimore: Lippincott Williams & Wilkins, 1998.)
Although skeletal muscle fibers look virtually identical on routine histologic stains (see Fig. 1-6), their biochemical differences can be appreciated with histochemical techniques. staining for the activity of myosin adenosine triphosphatase (ATPase), the enzyme that splits adenosine triphosphate (ATP) to yield energy for muscle contraction, reveals that some muscle fibers stain darkly (type II fibers), others lightly (type I fibers) (see Fig. 2-6). These histochemical results correlate with the physiologic properties of the muscle fibers themselves: type I fibers contract slowly (slow twitch) but can contract for long periods. Type II fibers contract quickly (fast twitch) but are more susceptible to fatigue. speed of contraction is therefore a property of the activity of myosin ATPase and the rate of ATP hydrolysis; endurance is related to the intracellular content and activity of mitochondria and the ability to generate ATP for contraction by oxidative or aerobic metabolism.
The specific types of muscle fibers that compose the muscle determine the functional characteristics of a whole muscle. Muscles that require sustained contraction, such as the antigravity muscles, typically contain more slow- twitch endurance fibers than do the muscles that contract briefly but quickly and with great force. it is even possible to show a difference among breeds of horses with regard to the muscle fiber composition of the same muscle; the middle gluteal muscle of the sprinting quarter horse is usually characterized by more of the large, fast-twitch, but low-endurance type ii fibers than is the same muscle of the slower but long-winded Arabian.
on casual examination with a light microscope, the cross-striations of skeletal muscle appear to be disks throughout the entire fiber. However, the electron microscope shows the striations only in the myofibrils and not in the sarcoplasm (cytoplasm of muscle cell). The alternate light and dark bands of all myofibrils appear at the corresponding places in the fiber (Fig. 8-2). The fact that corresponding bands of adjacent myofibrils are in register makes it appear that these bands extend across the whole fiber (Figs. 8-1 and 8-2). The apparent bands of myofibrils are due to relative density and partial overlapping of thick and thin filaments. Letters are used to designate the different bands.
Figure 8-2. Light micrograph (A) and electron micrograph (B) of longitudinal skeletal muscle and schematic representation (C) of a sarcomere. (Reprinted with permission of Wiley- Blackwell from Dellmann D. and Eurell J. Textbook of Veterinary Histology. 5th ed. Baltimore: Lippincott Williams & Wilkins, 1998.)
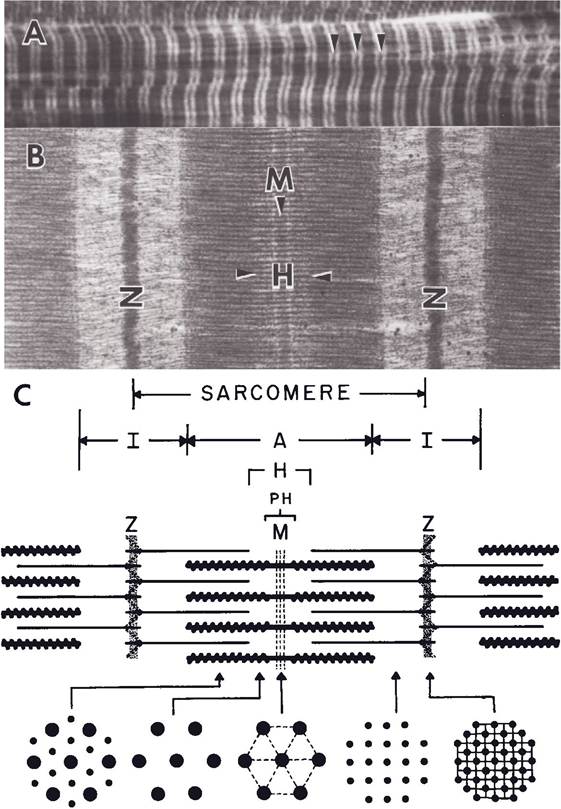
The light zones, or bands, called I bands, consist of thin filaments only. The darker regions, called A bands, are composed of overlapping thick and thin filaments (Fig. 8-2). Thus, alternating A and i bands produce the banding pattern on the myofibril.
A dense line, the Z line, bisects each I band (in fact, one end of each thin filament is attached to the Z line; the opposite end of each thin filament is free). The segment of myofibril between adjacent Z lines is the sarcomere, the fundamental unit of contraction in striated muscle (Fig. 8-2).
Each striated muscle fiber contains hundreds or thousands of myofibrils, and each myofibril contains approximately 1500 thick and 3000 thin myofilaments. Each thick filament is composed of hundreds of molecules of myosin, a golf club-shaped protein molecule with a molecular weight of 332,000 (by comparison, hydrogen has a molecular weight of 2). A thin filament is composed principally of chains of molecules of actin, a globular protein of molecular weight 70,000. The structure of these filaments and details of their actions in contraction are described in more detail in a later section.
The sarcoplasmic reticulum is agranular (contains no ribosomes) and functions in excitation-contraction coupling (discussed later in this chapter). A Golgi apparatus, large numbers of mitochondria, and glycogen inclusions also are found in muscle fibers.
The transverse tubules (or T systems) are continuous with the plasma membrane and extend into the interior of the muscle fiber at right angles in the myofibrils and sarcoplasmic reticulum. The T system propagates action potentials from the sarcolemma into the interior of the muscle fiber (Fig. 8-1) to initiate contraction of the entire fiber.
skeletal muscle fibers range in diameter from 10 to 100 μm. In general, the large fibers appear to be longer and tend to be found in large rather than small muscles. Animals on full feed are reported to have larger fibers than animals on restricted feed. It is generally accepted that males have larger muscle fibers than females. Length of skeletal muscle fibers varies widely with the length of the muscle and arrangement of muscle fibers (parallel or pennate). some fibers in parallel muscles probably extend the entire length of the muscle.
It is generally believed that skeletal muscle fibers are such specialized cells that little if any multiplication of fibers or formation of new fibers occurs after birth. All increases in size of muscles at any stage in life following birth are due to hypertrophy (increase in size) of individual muscle fibers, with the synthesis of more myofibrils and an increased vascular supply. It is well known that exercise can increase muscular development, as is seen in weight lifters. This, of course, is accomplished by increase in size of existing individual muscle fibers. If the nerve supply to a muscle is destroyed, the muscle fibers decrease to practically nothing, a condition called denervation or neurogenic atrophy. This was seen in sweeny of draft horses, when the suprascapular nerve was crushed by a collar, resulting in shrinking of the supraspinatus and infraspinatus muscles of the shoulder.
Excitation, Contraction, and Relaxation
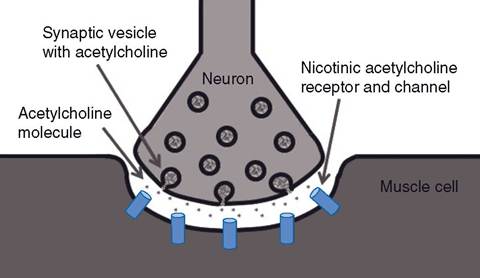
skeletal muscle contraction is triggered by the generation of an action potential on the sarco- lemma. This action potential is initiated by the firing of a motor neuron whose axon branch terminates at the neuromuscular junction near the midpoint of the muscle fiber. The neuromuscular junction (Fig. 8-3) is a type of excitatory synapse.
The action potential of the nerve is not propagated directly onto the adjacent muscle cell. Instead, depolarization of the motor nerve ending releases a chemical neurotransmitter, acetylcholine (ACh). When ACh diffuses across the synaptic cleft of the neuromuscular junction, it binds to specific cell membrane receptors (nicotinic ACh receptors) on the post-junctional membrane of the muscle fiber (Fig. 8-3). The binding of ACh to its receptors
Figure 8-3. Structural features of a neuromuscular junction. The terminal end of the axon contains synaptic vesicles with the neurotransmitter acetylcholine. Nicotinic acetylcholine receptors are on the cell membrane of the skeletal muscle cell.
brings about the opening of ligand-gated membrane channels, which allow sodium ions to enter the muscle cell.
The entrance of sodium (a positively charged cation) shifts the membrane electrical potential in a positive direction (depolarization). The effect of ACh persists only momentarily, since another enzyme, acetylcholinesterase, quickly degrades ACh.ACh is synthesized in the cytoplasm of the presynaptic nerve endings and stored in membrane-bound synaptic vesicles in the end of the nerve fiber (Fig. 8-3). Each action potential reaching the end of the nerve stimulates the release of a set number of vesicles and thus a set amount of ACh. Continuous synthesis maintains a constant supply of vesicles so that ACh is not depleted, even with intense exercise.
End-Plate and Action Potentials. The local depolarization of the sarcolemma at the neuromuscular junction is called an end-plate potential. This change in potential produces a local flow of current that depolarizes adjacent areas on the sarcolemma. Normally, the depolarization of the adjacent sarcolemma is enough to reach the threshold potential of electrically gated channels in these areas of the cell membrane. When this occurs, these channels open to allow the inward diffusion of sodium, and another action potential results. Local flow of current occurs again, and other action potentials are generated in other areas adjacent to the site of the first action potential. The process is repeated, and the overall effect is the propagation of action potentials over the entire sarco- lemma of the muscle fiber. This process is similar to that for propagation of action potentials along the axon of a nerve cell (see Fig. 2-16).
since the T tubules are inward continuations of the sarcolemma, the action potential (or impulse) travels along these tubules throughout the muscle fiber (Fig. 8-4). In certain sites where the T tubules pass immediately adjacent to the sarcoplasmic reticulum, there is a structural link between a protein in the sarcolemma of the T tubule (dihydropyridine receptor) and a membrane protein channel in the sarcoplasmic reticulum.
When an action potential occurs on the sarcolemma in the area of the dihydropyridine receptor, the channel in the sarcoplasmic reticulum becomes permeable to Ca2+. The permeable change is possible because of the link between the two membrane proteins.Before stimulation, the Ca2+ concentration within the sarcoplasmic reticulum is much greater (more than 100-fold) than within the sarcoplasm. When the Ca2+ channels in the sarcoplasmic reticulum open, Ca2+ diffuses into the surrounding sarcoplasm of the muscle fiber and into the myofibrils. The increase in Ca2+ in the myofibrils leads to the interaction of thick and thin filaments and movement (sliding) of the thin filaments past the thick filaments toward the center of the sarcomere. This sliding movement shortens the sarcomeres, which shortens the myofibrils, which shortens the entire muscle fiber. Figure 8-4 summarizes the movements of Ca2+ during excitationcontraction coupling in skeletal muscle.
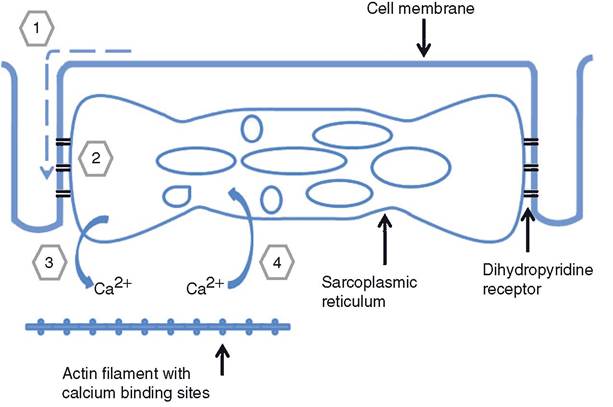
Figure 8-4. The sources and role of calcium during excitation-contraction coupling in skeletal muscle. An action potential is propagated along the T tubule (1). The action potential on the T tubule reaches a region with dihydropyridine receptors (2). Calcium ions are released from the sarcoplasmic reticulum to bind with regulatory proteins on actin filaments (3), and muscle contraction results. When calcium is released from its binding sites and transported back into the sarcoplasmic reticulum, muscle relaxation occurs (4).
Hyperkalemic periodic paralysis (HyPP) is an inherited disease of horses caused by a genetic mutation of a transmembrane protein. The electrically gated sodium channel (described earlier as participating in the generation of action potentials on the cell membranes of skeletal muscle) is defective in affected animals, and as a result, the permeability of the channel to sodium may be increased inappropriately. This permits the entrance of sodium, membrane depolarization, and involuntary muscular contractions. Hyperkalemia is an increase in serum potassium concentration, and this is one stimulus that can increase the permeability of the abnormal channels, hence the name of the condition. Clinical signs include muscle spasms, tremors, sweating, and weakness. This condition is also known as Impressive syndrome, because it is primarily seen in quarter horses and other descendants of the quarter horse sire Impressive.
Myosin and Actin Filaments. Each thick filament in a sarcomere is a bundle of myosin molecules. Each molecule has two parts: (1) a filament-like part that lies parallel to similar parts of other myosin molecules, making up the length of the thick filament, and (2) a part that projects outward like an arm from the end of the filament (Fig. 8-5). An enlargement at the end of the arm is termed the myosin head. The arm attaching the myosin head to the filament is flexible, like a hinge, where it joins the filament segment and also where it joins the head. Myosin heads protrude from all around the thick filament. They extend away from the center in both directions, toward the surrounding thin filaments (Fig. 8-5).
Each thin filament is made up of three proteins: actin, tropomyosin, and troponin. Actin molecules are the most prominent and are arranged in two long strands wound around each other in a spiral. Tropomyosin molecules are also joined in a strand that spirals around the strands of actin. The third protein, troponin, is found attached to tropomyosin at specific sites along the strand. Together they are called the troponin—tropomyosin complex (Fig. 8-5).
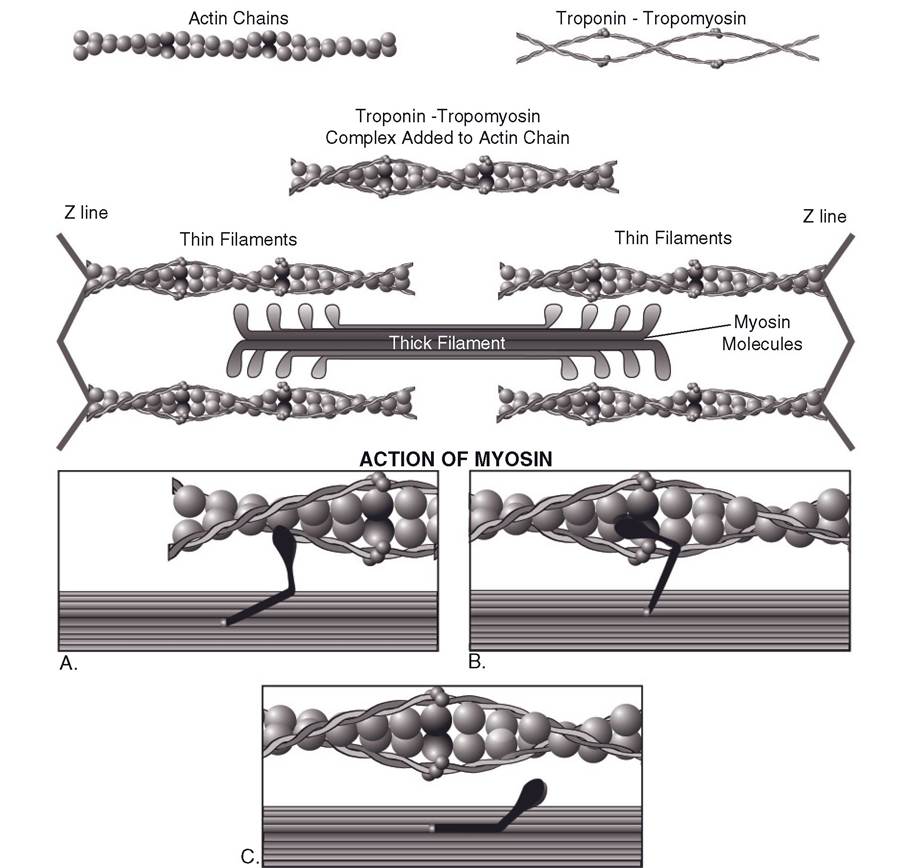
Figure 8-5. The primary components and the organization of thin and thick filaments are shown in the top-half of the figure. Boxes A, B, and C illustrate the sequential action of myosin as it binds to actin and then rotates to move the thin filament past the thick filament (from right to left in the figure).
The strands of tropomyosin lie over sites on the actin strand where myosin heads can bind. calcium ions released from the sarcoplasmic reticulum bind to the troponin part of the troponin-tropomyosin complex and induce a molecular change in the tropomyosin strand. This change uncovers myosin binding sites on the actin strands so that the myosin head can attach.
Binding of the myosin head to actin leads to the release of adenosine diphosphate (ADP) and phosphate, which were bound to the myosin head. The myosin head also rotates from its resting position toward the center of the sarcomere; this movement pulls the actin chain to which it is bound past the thick filament (Fig. 8-5).
The myosin head remains at its final angle and bound to the actin of the thin filament until an intact ATP molecule binds to another site on the myosin head. (This site on the myosin head is ATPase, which also promotes the hydrolysis of ATP prior to movement of the head.) With the binding of a new ATP, the myosin head detaches from the actin chain and resumes its resting angle. it is ready to repeat the process of attaching to actin, moving from resting to final angle and pulling the attached thin filament farther toward the center of the sarcomere, detaching, then binding still another ATP molecule.
The cycle of events that produces the shortening of each sarcomere, the sliding filament model of muscle contraction, is summarized in Figure 8-5. The resultant contraction shortens the sarcomere and shortens the I bands. The A band always remains the same length. During shortening, the thin filaments slide over the thick filaments as they are drawn from both ends, pulling the Z lines closer together (Fig. 8-2). The filaments themselves do not shorten. Relaxation. Muscle contraction will continue as long as there is an excess of Ca2+ in the sarcoplasm, but when the effect of the action potentials on the sarcolemma ends, the Ca2+ is sequestered back into the sarcoplasmic reticulum (Fig. 8-4). Ion pumps in the membrane of the sarcoplasmic reticulum use the energy of ATP to pump the Ca2+ from the sarcoplasmic fluid back into storage so that it can be ready for the next depolarization. (Without ATP the muscle cannot relax.) Only a small amount of Ca2+ is left out in the sarcoplasm of the relaxed resting muscle, not enough to act on the troponin-tropomyosin complex. Therefore, during relaxation the thin and thick filaments are dissociated, allowing the elasticity of the muscle to return it to its resting length, which pulls the Z lines and thin filaments back to their original positions.
Replenishment of ATP. A great amount of ATP is needed because the energy for contraction is derived from hydrolyzed (dephosphorylated) ATP. Also, the muscle fiber uses ATP to sequester Ca2+ back into the sarcoplasmic reticulum. It is also needed for complete recovery of the membrane after depolarization—the Na-K-ATPase system.
The concentration of ATP in resting skeletal muscle is relatively small, supplying only enough energy to maintain contraction for a brief period. since muscles continue to contract after the initial supply of ATP has been used, the resulting ADP is phosphorylated again from another source, creatine phosphate (CP).
There is normally about five times as much CP as ATP stored in the sarcoplasm of skeletal muscle. When the ATP is used for contraction and relaxation, transphosphorylation occurs from CP to the resulting ADP, forming ATP again. This replenishment reaction occurs almost as fast as ATP is used. Therefore, the ATP level changes little until the concentration of CP gets low. CP acts as the immediate energy source for the resynthesis of ATP. This is a convenient means of transferring energy for muscle contraction.
The concentration of CP is also limited. Thus, if muscle contraction continues for longer than a few seconds, the CP and new ATP eventually have to be reconstituted by the citric acid cycle (Krebs cycle) in the mitochondria of the muscle fibers. If muscle activity outstrips the ability of the mitochondria to produce ATP aerobically, anaerobic metabolism of carbohydrate fuel begins and lactic acid accumulates in the muscle cell. Glucose (the major carbohydrate fuel) is obtained from the blood supply to the muscle and from the glycogen stored in the muscle fibers. Glycogen is broken down by glycogenolysis. Glycogenolysis and glycolysis are complex processes involving a number of reactions, enzymes, and intermediate compounds.
oxygen from the blood must also be supplied to the mitochondria in muscles for the citric acid cycle to operate and result in oxidative phosphorylation of ADP to ATP. While lactic acid was mostly being produced anaerobically during muscle contraction, an oxygen debt was building up. This oxygen debt must
Table 8-1. Chain of Reactions That Supply Energy for Muscle Contraction and Recovery

ATP, adenosine triphosphate; ADP, adenosine diphosphate; CP, creatine phosphate.
be repaid during relaxation before optimal muscle activity can resume.
The chain of reactions involved in supplying energy for muscle contraction and recovery is shown in Table 8-1.
Strength of Contraction
Whenever a single muscle fiber receives a nervous impulse and action potentials are generated on the muscle fiber, the action potentials will be propagated over the entire fiber and cause the whole fiber to contract. This is the all-or-none law of muscle contraction.
The all-or-none law applies to a single muscle fiber or a single motor unit (a motor neuron and all of the muscle fibers it supplies); it does not apply to an entire muscle, such as the m. biceps brachii. The all-or-none law also does not state that a muscle fiber will always contract with the same speed or the same force; rather, it states that for the conditions at the time of stimulation, the muscle fiber will contract to its maximum. The force of contraction does depend on the state of the fiber at the time, that is, whether it is fatigued, stretched to its optimal length, and so on. The sliding filament mechanism is possible because of overlap of the thin and thick filaments. This overlap permits the binding between actin and myosin. Experimental studies have demonstrated that the amount of overlap of the filaments before contraction begins affects the contraction strength of individual muscle fibers. When muscle fibers are stretched before being stimulated, contraction strength increases up to an optimal amount of stretch. Any further stretching produces a decrease in contraction strength. This same relationship is true for the other striated muscle, cardiac muscle, and is an important factor in the regulation of cardiac contraction strength.
Summation. Each gross muscle, which is composed of multiple motor units and many individual muscle fibers, is capable of contracting with varying degrees of strength. This is the result of summing the contractions in two ways. Motor unit summation (recruitment) occurs when more motor units are stimulated to contract simultaneously in the gross muscle. Therefore, more muscle fibers and bundles are contracting and producing greater strength in the whole muscle. Temporal summation occurs when the frequency of stimulation to one or more motor units is increased. That is, the frequency of stimulation is such that the first contraction is not over by the time the second contraction begins. The two become additive, which increases the contraction strength (Fig. 8-6). Ordinarily, in normal muscle function both types of summation occur at the same time.
Tetany (Tetanus). When the frequency of stimulation becomes so rapid that further increases in frequency will not increase the strength of contraction, the greatest force that the muscle can develop has been reached. This is called tetany, tetanus, or tetanization. All of these terms are defined as a continuous tonic spasm of muscle or a steady state of contraction (Fig. 8-7). The disease caused by the toxin from Clostridium tetani, which produces spasm of the masseter muscles (lockjaw) followed by a spasm of other muscles, is also
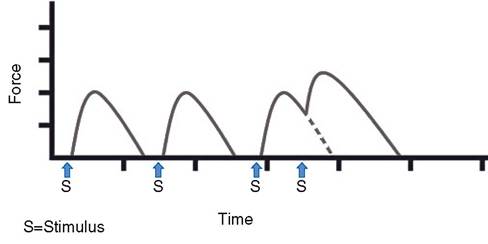
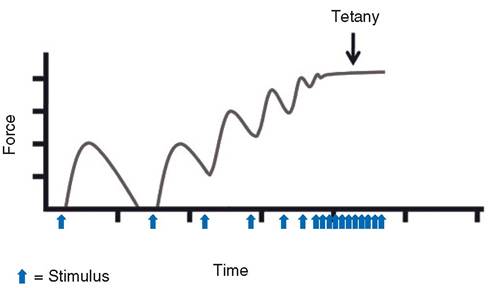
Figure 8-6. Temporal summation of muscle twitches. The first two contractions illustrate the force generated by a single stimulus (action potential). When a second stimulus is applied before the muscle can completely relax, summation occurs, and the force of contraction is greater (third recording).
Figure 8-7. Tetany. The interval between stimuli (action potentials) steadily decreases until no relaxation can occur between action potentials.
specifically referred to as tetanus. The Clostridium toxin produces skeletal muscle tetany by inhibiting the release of inhibitory neuromediators within the central nervous system. These inhibitors normally act within the spinal cord to regulate the activity of motor neurons to skeletal muscle. Without these inhibitors, any motor activity can result in spastic or tetanic contractions of the skeletal muscle. Loud noises or sudden movements can cause affected animals to tense their muscles and may induce generalized spasms.
Fatigue. Fatigue is a decrease in work capacity caused by work itself. Fatigue may occur at the level of the individual muscle fiber, or it may be a generalized state affecting the animal as a whole. on a whole-animal basis, resistance to fatigue entails some poorly defined factors such as motivation. The factors contributing to fatigue of an individual muscle fiber have been best studied in muscle cells in isolated in vitro preparations outside of the body. These studies indicate that fatigue is a function of the muscle cell itself and is not due to failure or fatigue of the neurons that innervate skeletal muscles.
At the individual cell level, one contributor to fatigue is a decrease in the availability of ATP, as its use increases with increased muscle contractions. However, even with strenuous prolonged exercise ATP is never totally depleted, because the intracellular generation of ATP is also increased during exercise. increases in intracellular concentrations of various metabolites generated as a result of contraction also contribute to fatigue. Among these are the phosphate from the splitting of ATP to ADP and organic acids such as lactic acid. Lactic acid can diffuse out of the cell and reduce the pH of the interstitial fluids within a muscle. Accumulations of lactic acid within muscles contribute to pain and soreness of muscles following strenuous exercise.
The rate at which ATP is generated within an exercising muscle depends on the availability of substrates for the different metabolic pathways that produce ATP. If a muscle primarily uses the glycolytic pathway, glucose must be readily available. Glucose may be stored within the muscle cell as glycogen or delivered via the circulation. if a muscle uses the oxidative pathway to produce ATP, oxygen must be available in addition to fatty acids or other similar substrates. The oxygen, of course, must arrive via the circulation. Adequate blood flow during exercise to deliver oxygen, fatty acids, and/or glucose to skeletal muscle is a key factor in resisting fatigue.
Drugs That Affect Skeletal Muscle Function
Almost as soon as ACh initiates the impulse for muscle contraction, the enzyme acetylcholinesterase inactivates the ACh. This enzyme, which degrades ACh, is found within the area of the neuromuscular junction. Agents that are capable of inhibiting the action of the acetylcholinesterase enzyme are referred to as anticholinesterases. The effect of anticholinesterases at the neuromuscular junction is to prolong the availability and effects of the ACh.
These actions may be desirable in some disease states, and neostigmine and physostigmine are commonly used anticholinesterase drugs. However, too much anticholinesterase activity can produce a toxicity characterized by muscular spasms and asphyxiation due to spasms of skeletal muscles necessary for respiration. The signs of anticholinesterase toxicity also include constriction of the pupil of the eye, intestinal cramps, vomiting, and diarrhea. These signs are due to the increased availability of ACh at neural synapses within the parasympathetic division of the autonomic nervous system. Parasympathetic nerves are discussed in more detail in Chapter 10.
Anticholinesterase activity is the basis for some effective insecticides and the so-called nerve gases that have been studied extensively by the armed forces since World War
II. Most of these compounds are alkyl phosphates, and the insecticides are also known as organic phosphates or organophosphates. The insecticides include such products as malathion, parathion, and diazinon. Products for both external application and oral administration are included in the organophosphate insecticides. If improperly used, any of the organophosphates is extremely dangerous, not only to domestic animals but also to the person using it. Therefore, it is imperative that this class of insecticides be used under proper supervision and that instructions for use are followed exactly.
Another group of drugs affecting the neuromuscular junction are the curariform drugs. These drugs act like curare, the deadly poison that some South American Indians use on their arrowheads. These agents bind to ACh receptors on the postjunctional membrane so that ACh cannot bind to the receptors and produce an end-plate potential. Curare is not destroyed by acetylcholinesterase. Death can result from asphyxiation because of paralysis of muscles needed for respiration. The paralysis induced by curariform drugs depends on the concentration of the drug, and such agents are used therapeutically to produce varying amounts of paralysis, such as for muscle relaxation during certain types of surgery.
Botulinum toxin, which is produced by the bacterium Clostridium botulinum, also acts at the neuromuscular junction. It prevents the release of ACh from the nerve ending by preventing synaptic vesicles containing ACh from binding to the cell membrane for exo- cytosis. Flaccid paralysis results because action potentials cannot be produced on the sarcolemma to bring about muscle contraction. The term limberneck has been applied to the characteristic flaccid paralysis of the neck in poultry intoxicated with botulinum toxin.
Types of Muscle Contraction
The primary function of muscle is to contract, that is, develop tension and shorten. However, contraction is often described as having four types: concentric (shortening), eccentric (lengthening), isometric (same length but increased tension), and isotonic (same tension but length changes).
Concentric contraction is the usual form, in which the muscle moves a bone or segment by shortening. An example is flexion of the elbow by contraction of the m. biceps brachii.
Isometric contraction occurs naturally whenever a limb or portion of the body is held stationary against equal resistance, such as gravity. To hold the head up in a fixed position, the dorsal neck muscles must contract isometrically.
Eccentric contraction occurs in the extensor muscles of the neck when an animal lowers its head gradually. Antagonistic muscles may also undergo eccentric contraction when unsuccessfully opposing the actions of a prime mover.
Isotonic contraction occurs when the length of the muscle changes but the tension remains the same, primarily when a muscle lifts a given weight. The weight is constant; therefore, the tension does not change.
Rigor and Rigor Mortis. If most of the ATP in a muscle is depleted, the myosin heads cannot separate from the actin in the thin filaments and the calcium can no longer be sequestered back into the sarcoplasmic reticulum by the calcium pump. Therefore, relaxation cannot occur because the actin and myosin filaments are bound in a continuous contracted state. This is termed rigor, and it is sustained until more ATP is made available. Rigor mortis is rigor that occurs a few hours after death, when ATP is no longer available. The muscles of the whole animal progressively become stiff and rigid; without ATP to sequester the calcium and separate the cross-bridges, the filaments remain locked together. The rigidity continues until cell autolysis and protein degradation break down the muscle.
Tone. The term muscle tone refers to the slight tension exhibited by all muscles at rest. it is due to the continuous transmission of impulses at very low frequency from the spinal cord to the muscles. Tone keeps muscles in a partially contracted state and prevents them from becoming flaccid (flabby), as occurs in paralysis.
When an animal becomes anxious, fearful, or excited, the muscle tone intensifies. Therefore, the muscles become taut (tension increases) and the animal can respond faster to any stimulus. This is often seen in the skittish, nervous, or jumpy animal. During sleep, muscle tone is low to allow for optimal relaxation.