Smooth Muscle
Smooth muscle is sometimes called involuntary muscle because it is found in structures that are not voluntarily or consciously regulated (e.g., gastrointestinal tract, blood vessels, reproductive tract).
About 99% of the smooth muscle of the animal body is of the visceral type, which is also called single-unit or unitary smooth muscle. Cells of this type of smooth muscle are joined with gap junctions to provide mechanical and electrical connections between cells. The electrical connections permit the propagation of action potentials directly from cell to cell. The spreading of action potentials and the contraction that is elicited allow a group of muscle cells to act together as a single unit. This type of activity is appropriate for generalized activities of an entire organ, such as the contraction of the stomach.About 1% of the smooth muscle of the animal body is multiunit smooth muscle (e.g., iris and ciliary body of the eye and pilomotor fibers that erect the hair in the skin). in this type of smooth muscle, the contraction of each individual smooth muscle cell is more dependent on its autonomic innervation. Gap junctions are not prevalent between these cells.
Structure
The smooth muscle cell is a fusiform (spindleshaped) contractile unit with a central nucleus. size of smooth muscle fibers varies considerably. Most cells are 50-250 μm long and 510 μm in greatest diameter. The major portion of the cell consists of sarcoplasm. No crossstriations, myofibrils, or sarcolemma are easily visible with the light microscope (see Fig. 1-6). Filaments are present as actin and myosin molecules, but there are no orderly arrangements to form striations. interactions between actin and myosin filaments (i.e., sliding filaments) are believed to be the basis for smooth muscle contractions, but the characteristic shortening of sarcomeres is not seen because there is no distinct organization of the filaments.
A sarcoplasmic reticulum, which accumulates calcium, is present, but it is not as extensive or as highly organized as in skeletal muscle.Like skeletal muscle, smooth muscle cells can hypertrophy to increase the size of organs. However, unlike skeletal muscle, smooth muscle cells can also divide mitotically to increase the number of cells. For example, the increase in size of the uterine wall during pregnancy to several times its nonpregnant volume is due in considerable measure to an increase in the amount of smooth muscle in the wall. some of the increase is due to an increase in size of individual muscle fibers, but there is also an increase in the number of cells. This hypertrophy and hyperplasia in the uterus are under the influence of reproductive hormones. Late in pregnancy there is also a change in the structure of the cell membranes of uterine smooth muscle cells. An increase in gap junctions between cells in late pregnancy sets the stage for the mechanical and electrical coupling between smooth muscle cells that will be necessary for normal parturition. in general, the structure and function of smooth muscle are more subject to modification by external factors, such as hormones, than are those of skeletal muscle.
Stress-Relaxation
smooth muscle exhibits a special property called stress-relaxation, or plasticity. This is the ability to adjust to stretching without increasing the final tension or the pressure exerted on the contents within a hollow viscus surrounded by smooth muscle. As the muscle stretches, the tension increases at first, but then in a few seconds or a few minutes the smooth muscle relaxes again to its original tension, even though it is still elongated.
This stress-relaxation occurs in the stomach when it is filling with food, in the intestines as the processed food moves along, in the blood vessels when the blood volume increases, in the urinary bladder as it increases its volume of urine, and in the uterus as pregnancy develops.
Plasticity allows expansion of stretch within physiologic limits without an increase in pressure and without pain; the smooth muscle does not lose its contractile ability. The reverse occurs upon emptying of a visceral organ when the stretched muscle shortens back to its original length: all tension is lost at first but returns shortly. Plasticity is believed to be due to changes in the arrangement or binding of the myosin and actin filaments upon stretching or shortening.
Contraction and Relaxation
All skeletal muscle contraction depends on Ach release at a neuromuscular junction and the generation and propagation of action potentials on the cell membrane. This is not the case for smooth muscle. The stimuli that bring about smooth muscle contraction and relaxation are quite variable. Thus, smooth muscle is a much more functionally diverse tissue than skeletal muscle.
The contraction and relaxation of most smooth muscle are much slower events than the rapid muscle twitch that is characteristic of skeletal muscle. This permits the maintenance of a relatively constant pressure with the use of little cell energy. This is characteristic of smooth muscle in organs that require a constant state of tone or some degree of contraction for normal function (e.g., smooth muscle in the walls of blood vessels).
Role and Sources of Calcium
As in skeletal muscle, contraction and relaxation of smooth muscle are linked to the Ca2+ concentration in the cytosol of smooth muscle cells. However, how this concentration is regulated and the role of Ca2+ in the contraction process is quite different in the two types of muscle. some calcium is stored in the sarcoplasmic reticulum of smooth muscle cells, but many types of smooth muscle cells also contain a significant number of calcium channels in the outer cell membrane (Fig. 8-8). These calcium channels may be either voltage-gated or ligandgated, and the smooth muscle within a given organ may have both types of channels or primarily only one type of channel.
When these channels open in response to the appropriate stimulus (changes in membrane potential or presence of specific ligands), calcium can diffuse into the cell to initiate contraction (Fig. 8-8). Receptors that are specific for many different ligands (e.g., hormones and neurotransmitters) are found on the smooth muscle of different organs. This is important functionally, because a given hormone produces contraction only of organs with receptors for that specific hormone. Calcium entering from the outside via the channels may stimulate the release of further calcium from the sarcoplasmic reticulum, which further strengthens the contractions.An understanding of the role of calcium channels in the outer cell membrane of smooth muscle cells led to the development of a group of drugs termed calcium channel blockers. These agents are capable of binding to and inactivating these calcium channels, and this tends to reduce the strength of contractions. Smooth muscle cells in the walls of many arterial blood vessels have these types of channels, and this type of drug has been proved effective in lowering blood pressure in both humans and animals.
When cytosolic Ca2+ increases within smooth muscles, calcium ions bind to a regulatory protein (calmodulin) (Fig.8-8). Unlike skeletal muscle, whose regulatory proteins are associated with the actin filaments, the calmodulin in smooth muscle is associated with the heads of myosin molecules in myosin filaments. The binding of calcium to calmodulin activates a kinase, also associated with myosin, and this kinase phosphorylates other sites on myosin. This phosphorylation ultimately results in contraction, so myosin phosphorylation is a key regulatory step (Fig. 8-8). This is in contrast to skeletal muscle, whose myosin does not require phosphorylation. For relaxation, the phosphorylation of myosin is reversed by other intracellular enzymes that are always present and active (Fig. 8-8). This reversal and relaxation can occur when the cytosolic calcium concentration is reduced.
Calcium is removed from the cytosol by the transport back into the sarcoplasmic reticulum or out of the cell. Most of this transport is done by an active transport system (Ca2+-ATPase) on the membrane of the endoplasmic reticulum and the outer cell membrane.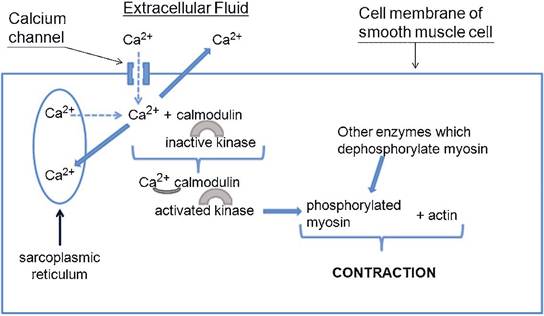
Figure 8-8. Sources and role of calcium ions in the initiation and regulation of smooth muscle contraction. Dotted lines associated with calcium ions indicate movement by diffusion. solid lines associated with calcium ions indicate removal of calcium from the cytosol to promote relaxation. Relaxation can also be promoted by the action of other intracellular enzymes that dephosphorylate myosin.
Unlike skeletal muscle, some smooth muscle responds to certain stimuli by relaxation or a reduction in contraction strength. For example, stimulation of β2-adrenergic receptors on airway smooth muscle in the lungs produces relaxation and an increase in airway diameter. in general, this type of relaxation is due to either a reduction in the number or availability of calcium channels in the cell membrane or a reduction in the phosphorylation of myosin within the myofilaments of smooth muscle cells.
Action Potentials and Slow Waves
Not all smooth muscle cells have action potentials on their cell membranes during contraction and relaxation. cells with predominantly ligand-gated calcium channels in their cell membranes may undergo a contraction-relaxation cycle without action potentials. The cell membrane may depolarize slightly because of the entry of calcium, but no action potential is seen. smooth muscle cells that do have action potentials during contraction have both sodium and calcium electrically gated channels in their cell membrane.
Action potentials in skeletal muscle occur after the binding of Ach to its cell membrane receptors. in smooth muscle, action potentials can be elicited by a variety of stimuli.
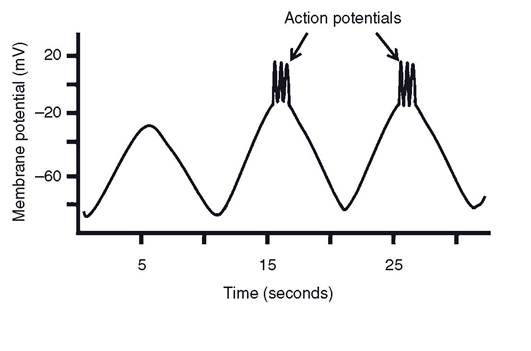
some smooth muscle is similar to skeletal muscle in that action potentials occur only after the binding of ligands to cell membrane receptors, but action potentials may also occur in smooth muscle in response to mechanical stretch or during slow-wave electrical activity. The smooth muscle that responds to stretch is believed to have membrane channels that are subject to mechanical stimulation.The visceral or single-unit smooth muscle in some organs (the gastrointestinal tract being the classic example) exhibit a unique type of membrane electrical activity termed slow waves. These are waves, or periods, of spontaneous fluctuations in the resting membrane potential that spread throughout a body of smooth muscle and typically occur at some regular rhythm (Fig. 8-9). Slow waves alone do not cause contractions, but action potentials may occur at the peak of these waves, and the action potentials are associated with contractions (Fig. 8-9). Because action potentials and contractions are seen only at the peak of the slow waves, the rate at which slow waves develop determines the rate at which smooth muscle contractions can occur. The precise origin of the slow waves is uncertain, but a variety of stimuli, such as hormones, neurotransmitters, and the local chemical environment, can determine whether action potentials occur at the peak of the slow waves. The relationship between slow-wave activity and action potentials is important in the regulation of gastrointestinal motility and is discussed in more detail in Chapter 21.
Action potentials spread across groups of single-unit smooth muscle fibers because of gap junctions between the fibers (where the plasma membranes of adjacent cells touch). Thus, single-unit smooth muscle cells can be linked electrically while remaining independent chemically (no secretion of transmitter substance from cell to cell is required).
Figure 8-9. Slow-wave electrical activity in smooth muscle with action potentials occurring at the peak of slow waves.
Autonomic Innervation
In visceral or single-unit smooth muscle, the fibers of the autonomic nervous system travel between the smooth muscle cells in a branching network of terminal fibrils. These fibrils have varicosities (beadlike enlargements) at intervals along their axons. When action potentials depolarize them, the transmitter substance is released and diffuses to the smooth muscle cell membranes of several cells, where stimulation occurs. in multiunit smooth muscle, a branch of an autonomic nerve innervates each muscle fiber. This provides more direct neural control of each muscle cell, but the junction between neuron and muscle is less highly structured than the neuromuscular junction of skeletal muscle. For both types of smooth muscle, the innervation is usually dual; that is, both divisions of the autonomic nervous system innervate smooth muscle. important exceptions are blood vessels (arteries, arterioles, and veins), which have predominantly sympathetic innervation, and in the skin, where the pilomotor fibers and sweat glands receive only sympathetic innervation.
Acetylcholine is released from the parasympathetic postganglionic nerve fibers and norepinephrine from the sympathetic postganglionic fibers. The response of smooth muscle (contraction or relaxation) to these neuromediators depends on the type of autonomic receptor (see Tables 10-1 and 10-2) on the smooth muscle and the intracellular events initiated by the binding of the neuromediators to their receptors. For example, stimulation of β2-adrenergic receptors causes smooth muscle relaxation, while stimulation of α1-adrenergic receptors causes smooth muscle contraction. some smooth muscle can also be stimulated to contract by stimulation of muscarinic receptors by ACh. individual smooth muscle cells may have multiple types of receptors in their cell membranes and respond to both autonomic neurotransmitters. in this case, the overall response of the smooth muscle also depends on the relative levels of the different neurotransmitters.