Spinal reflexes
Key points
■ Reflexes are hard-wired into the nervous system and most are polysynaptic.
■ Reflexes can be ipsilateral, contralateral, intrasegmental or intersegmental, somatic or autonomic.
■ Muscle spindles detect stretching of muscle. They provide sensory input to the CNS using 1a afferent fibres and their intrafusal fibres receive motor input from the CNS via γ-efferent fibres. α-LMN innervate the extrafusal muscle fibres.
■ The myotatic (‘stretch’) reflex has a significant role in maintaining body posture. Gravity causes muscle stretching, stimulating muscle spindles that induce reflex contraction of the muscle.
■ The Golgi tendon organ inverse myotatic reflex protects the skeletal muscle from excessive contraction by causing reflex muscle relaxation.
■ Muscle tone is controlled by activity of α and γ-LMN; these LMNs are stimulated by muscle spindles and specific UMN tracts.
A reflex is defined as a stereotypical response produced by a specific stimulus. The reflex arc involves receptors and nerves of the PNS (sensory and motor) and a region of the CNS in which the sensory input connects to the motor output. This CNS region is in the brain for cranial nerve reflexes and in the spinal cord for limb and body reflexes. Reflexes are ‘hard-wired’, meaning that the neuronal connections are established during embryonic development and are present at birth. Conversely, neural responses (e.g. the menace response) have to be learned. Their ‘wiring’ develops postnatally as a consequence of experience. The reflex is the functional unit of the nervous system as compared with the morphological unit that is the neuron.
That the reflex is the most basic functional unit of the nervous system is evidenced by its presence in simple metazoan animals. In vertebrates, incoming sensory input into the dorsal horn can synapse directly with LMNs in the ventral horn of that spinal cord segment and stimulate a motor output.
This simplest reflex is called a monosynaptic reflex and is exemplified by the patellar reflex. Most other reflexes involve interneurons interposed between the input and output side. Interneurons permit divergence (see Fig. 4.9) so that the input can be distributed to a wider population of output neurons. Thus one type of input can stimulate both agonist muscle contraction and antagonist muscle relaxation, e.g. the patellar reflex can stimulate quadriceps muscle contraction and hamstring muscle relaxation (see Fig. 4.4).There are a number of different types of reflexes including:
(a) Somatic reflexes that permit the animal to respond to the external environment. An example of this is the palpebral reflex, where stimulation around the eyelids causes blinking to protect the eyeball.
(b) Autonomic reflexes that permit the animal to respond to changes in its internal environment. An example of this is the increase in heart rate in response to reduced blood pressure.
Reflex arcs can be ipsilateral, contralateral due to decussation of the fibres, intrasegmental located within one spinal cord segment, or intersegmental involving a number of different spinal cord segments.
Muscle spindles
Muscle spindles are spindle/fusiform-shaped receptors located within striated/skeletal muscles. They are called stretch receptors as they detect stretching of the muscle. Muscle spindles comprise modified muscle cells called intrafusal fibres that have contractile elements at the ends of the spindle and sensory receptors in the middle. The surrounding normal muscle fibres are referred to as extrafusal. Muscle spindles are located in parallel with the extrafusal muscle fibres and, consequently, are stretched when the muscle is stretched. The contractile elements of muscle spindles are innervated by γ-motor neurons that, when stimulated, cause contraction at the ends of the spindle, thus stretching the centre of the spindle. The intrafusal fibres and the γ-motor neuron comprise the fusimotor system.
Stretching of the muscle spindle causes firing of the sensory receptors in the spindle and stimulation of the 1a afferent nerve fibres. Sensory impulses travel via the spinal nerve to the spinal cord, or, if arising from head muscles, via cranial nerves to the brainstem. In the CNS the 1a afferent fibre synapses with α-LMNs resulting in stimulation of extrafusal fibres and muscle contraction. The extrafusal fibre plus the α-LMN is the skeletomotor system (Fig. 5.1).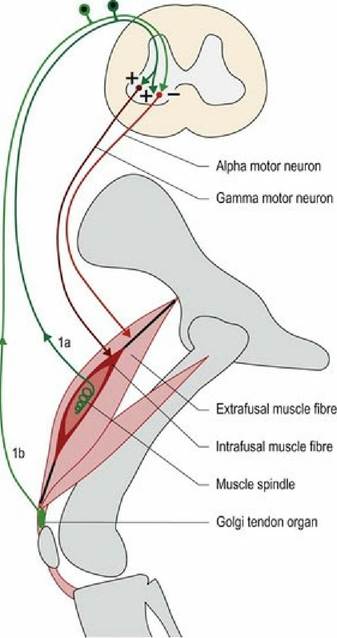
F⅛. 5.1 Muscle spindles are in parallel with the extrafusal fibres and are stretched when the muscle is stretched. Golgi tendon organs are in series with the extrafusal fibres and are stretched when the muscle shortens due to contraction.
The myotatic reflex
The myotatic (‘stretch’) reflex arc is a feature of limb and trunk muscles especially. It uses the stimulus of muscle stretching to generate nerve impulses in the 1a afferent fibre. The impulses travel to the spinal cord, synapse (usually via interneurons) onto the α-LMN supplying that same muscle, stimulating nerve impulses in that α-LMN. This causes contraction of the extrafusal fibres surrounding the muscle spindle, thereby shortening the muscle and reducing the stretch of the muscle spindle. Thus, activity in the 1a afferent nerve fibre stimulates agonist muscle contraction. Simultaneously, it also causes reciprocal inhibition; that is, it inhibits contraction in the antagonist muscle. For example, the effect of gravity on the weight-bearing stifle joint is to make it collapse (stifle flexion), thereby stretching the quadriceps muscle and its muscle spindles. Spindle stretching causes reflex contraction of the quadriceps muscle and stifle extension to support the animal’s weight. Simultaneously, the stifle flexors (hamstring muscles such as the semimembranosus and semitendinosus muscles) will be inhibited. The input from the 1a afferent is also forwarded to the brain to provide proprioceptive information that is essential for planned and coordinated motor function (see Chapter 6).
Different types of 1a fibres are stimulated by static versus dynamic stretching. Stretching of the muscle spindle and stimulation of 1a afferents can be caused by the following.
(a) The effect of gravity causing the joint to flex and passively stretching the extensor muscles. This causes reflex contraction of α-LMN and is used to sustain posture (see ‘Posture and the myotatic reflex’ in this chapter).
(b) Stretching of tendon and hence the muscle, by tapping of the tendon. For example, the patellar reflex is elicited clinically by tapping the patellar ligament (the continuation of the quadriceps tendon) with a patella hammer, or plexor (see Fig. 4.4 and 13.9).
(c) By activation of γ-LMNs causing intrafusal muscle fibres to contract. This is the mechanism by which many descending UMNs ultimately cause contraction in the muscles - they stimulate the γ-motor neuron, which reflexively causes stimulation of the α-LMNs and amplification of the action.
Within the spinal cord the input from the 1a afferent fibres can link to LMNs in the same spinal cord segment or in different spinal cord segments, ipsi- or contralaterally (see Fig. 4.8). The propriospinal tract is a white matter tract immediately surrounding the grey matter of the spinal cord, and conveys axons connecting between spinal cord segments. Thus input from one muscle can influence other muscles acting around that joint, or other joints in the same limb, or other limbs. For example, in the withdrawal reflex, noxious stimulation of the foot causes limb flexion. If the animal is standing at the time, then the other limbs will extend to compensate for the loss of weight-bearing in the stimulated limb. The withdrawal reflex uses multiple spinal cord segments to activate both multiple flexor muscles within the stimulated limb, and extensor muscles of the other limbs.
Golgi tendon organs and the inverse myotatic reflex
Golgi tendon organs are located within the tendon; therefore they are in series with the muscle.
They are stimulated by the stretching when extrafusal muscle fibres contract, causing muscle shortening and increased tension in the tendon.Their sensory innervation is type 1b afferent fibre; this has a higher threshold to stimulation than the muscle spindle. Significant stretching of the tendon stimulates nerve impulses in 1b afferent fibre; these are projected into the spinal cord. This input stimulates interneurons that inhibit the α-LMN of that muscle, thereby decreasing muscle contraction and reducing the tension in the tendon (see Fig. 5.1). It may also stimulate α-LMNs of the antagonist muscle.
The activity of the Golgi tendon organs functions to prevent over-contraction of the muscle and muscular avulsion (tearing of the muscle fibres or the tendon) from the bone. It also facilitates switching from extensor to flexor activity.
α-γ co-activation
When the α-LMNs are stimulated, the extrafusal fibres contract, reducing the stretch of the intrafusal fibres; this decreases activation of the Ia afferent fibre. This decreased sensory input to the CNS would result in loss of proprioceptive input (Chapter 6). But if the γ-LMN is activated simultaneously with the α-LMN, then the intrafusal fibres contract, causing comparable shortening of the muscle spindle. Thus the relative stretch of the muscle spindle is maintained and the Ia firing is sustained. Therefore when a muscle is stimulated to contract, both α- and γ-LMNs are simultaneously stimulated and proprioceptive input from the muscle about its length and tension is maintained. This is called α-γ co-activation. Functionally, this acts to maintain appropriate muscle tone, despite changes in muscle length, and to maintain continual proprioceptive input to brain, which is essential for normal posture and movement.
Muscle tone
Muscle tone refers to the muscle’s resistance to being stretched. Muscle tone is due to several factors including the tonic γ-loop mechanism.
Constant activation of the γ-efferents causes low-level stimulation of the Ia afferents, which causes low-level stimulation of the α-LMN and low-level contraction of the extrafusal fibres. This manifests as muscle tone and resistance of the muscle to being stretched. The inherent elasticity in muscle components also contributes to tone. Tone is further enhanced by the effect of specific UMN tracts, facilitating LMNs (α and γ). Clinically, loss of LMN output to muscle will result in loss of tone. Loss of LMNs occurs with either damage to the cell body in the CNS or the axon in the PNS.Limb muscle tone can be assessed by laying the animal on its side and gently flexing and extending the limbs. Limb flexion will cause stretching of the extensor muscles and extension will cause stretching of the flexor muscles; both activities induce myotatic reflex-induced contraction in stretched muscles. The reflexively induced contraction will cause resistance to flexion and extension, respectively. Assessment of limb muscle tone is an important component of the neurological examination and can help differentiate UMN from LMN signs, thus aiding lesion localisation.
Posture and the myotatic reflex
The myotatic reflex is reflex activity based on muscle spindle input. It is a major mechanism by which the animal maintains posture and supports itself against gravity. Gravity acts on the animal to cause flexion/collapse of the limb joints, flexion of the cervical vertebral column due to the weight of the head and extension (lordosis) of the thoracolumbar vertebral column due to the weight of the trunk. Thus gravity causes stretching of the muscle spindles in the extensor muscles of the limbs and neck, and flexor muscles of the thoracolumbar vertebral column. The stretching causes firing of the 1a sensory fibres from the spindles, input via the spinal nerves to the spinal cord, stimulation of the α-LMNs and extension of the limbs and neck and flexion of the vertebral column. Simultaneously, the antagonistic muscles (limb and neck flexors and spinal extensors) may be inhibited (Fig. 5.2).
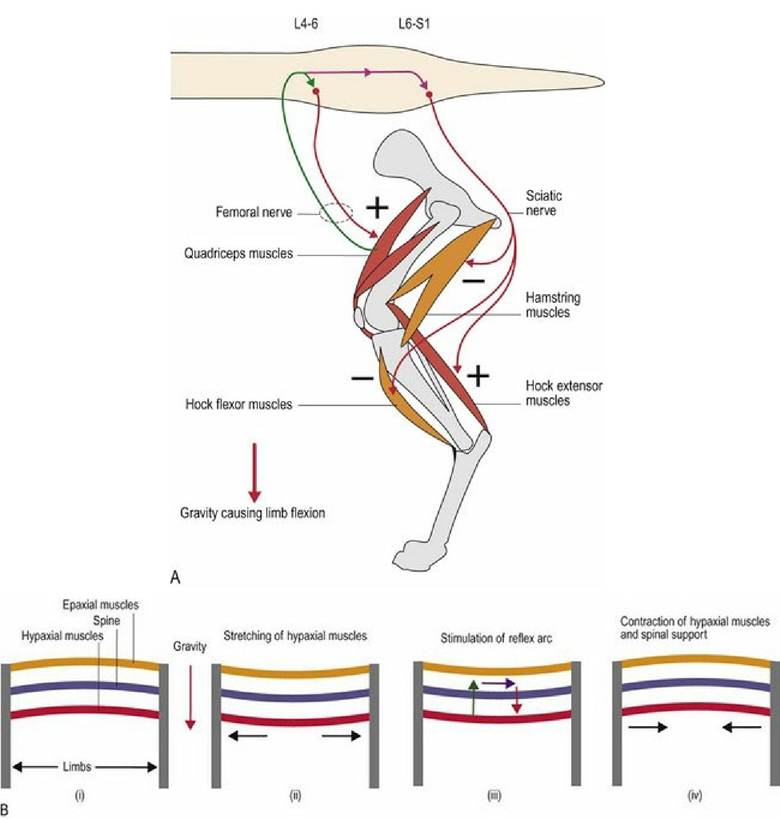
Fig. 5.2 The myotatic reflex and postural support. The effect of gravity on (A) the pelvic limb and (B) the vertebral column. The quadriceps and hock extensor muscles (A) and hypaxial muscles (B) will be stretched
by the effect of gravity, resulting in reflexively stimulated contraction and postural support against the effects of gravity.
Posture is also supported passively by elastic ligaments. For example, the nuchal ligament extending from the dorsal spinous processes of the cranial thoracic vertebrae to cervical vertebrae is analogous to a gigantic rubber band. It is stretched when the head is lowered (neck flexion) and recoils to help raise the head, acting to decrease the work required by the neck extensor muscles.
Reflexes in different parts of the body
Clinically, reflexes are tested to evaluate the integrity of the different components of the reflex arc. This includes the sensory input (receptor and spinal or cranial nerve), the CNS region in which input connects to LMN cell body (brain and spinal cord), and the motor output (spinal or cranial nerve, neuromuscular junction and muscle). Disruption of any one of these three components will result in decreased, or loss of, reflex activity. This is most commonly seen with lesions that involve either the region of the CNS in which the LMN cell body is housed, or in the LMN itself. Such a lesion is called a lower motor neuron lesion as it damages the LMN cell body or axon. If the reflex arc is present then the three anatomical components are intact and functioning.
Clinically, observing that a reflex is decreased, can particularly help localise a lesion. The clinician can identify which specific regions of the PNS and the interconnecting CNS have been compromised. For example, loss of the patellar reflex in the dog, means that the femoral nerve (afferent and efferent components of the reflex arc), or spinal cord segments L4-L6 have been compromised. Conversely, an intact reflex arc tells the clinician that the lesion does not involve that area of the CNS or the PNS. This is useful, but does not localise the lesion within the rest of the CNS.
It should be noted that the UMN system has a significant, inhibiting influence on LMNs especially those LMNs supplying extensor (antigravity) muscles. A CNS lesion that is cranial to the area of the CNS involved in the reflex arc may block UMN inhibition of the LMN of the reflex arc. This lesion damages the upper motor neurons, and hence is called an upper motor neuron lesion. Due to loss of the inhibitory influence of the UMN, the reflex may be exaggerated. This exaggerated reflex can indicate the presence of a lesion in the UMN system cranial to the location of the reflex wiring. For example, if there is a lesion in the thoracolumbar spinal cord, then the patellar reflex will be intact and may even be exaggerated.
Clinically, there are a number of reflexes that can be evaluated in the animal. They include limb, trunk and head reflexes and are described in Chapter 13.