Structural and functional anatomy
The vestibular system is divided into peripheral and central components (Fig. 8.1); this segregation has clinical implications. The peripheral vestibular system consists of the vestibular components of the inner ear (receptors and axons of CN VIII), while the central vestibular system comprises the vestibular nuclei and their output.
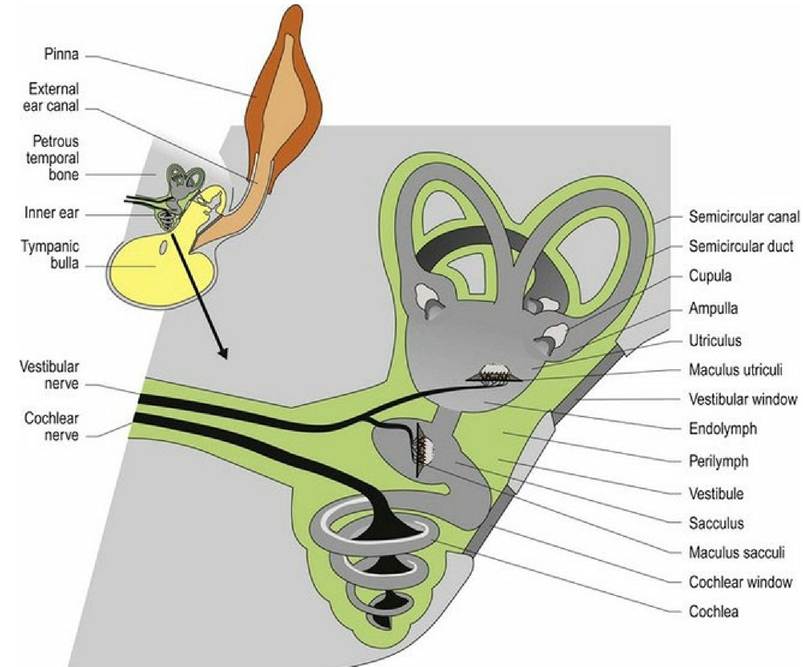
Fig. 8.1 The vestibular system of the inner ear.
The inner ear is housed in the petrous temporal bone bilaterally. It contains a number of canals and cavities forming the bony labyrinth. On each side there are three semicircular canals, the bases of which connect to the bony vestibule. Ventral to the vestibule is the spiral cochlear canal that houses the auditory receptors (see Chapter 10). The semicircular canals arise from the vestibule, curve around and connect back to the vestibule. At one end of those connections the canal is dilated to form the ampulla. The canals and cavities are lined with membranes forming the membranous labyrinth. Perilymph is the fluid located between the membrane and the bone while endolymph fills the membranous labyrinth.
Static position, linear acceleration and deceleration of the head
Located within the vestibule are expanded portions of the labyrinth, called the utriculus and the sacculus (Fig. 8.1). These membrane-bound portions of the vestibular system each have a region called a macula; this is where the hair cells are located. The stereocilia and kinocilium of each cell project into an overlying gelatinous matrix called the otolithic membrane. Embedded in the membrane are calcareous crystals called otoliths that, having sufficient mass, move under the effect of gravity, pulling on the otolithic membrane and causing deflection of the stereocilia and kinocilium. The macula in the utriculus is oriented in the horizontal plane, whereas in the sacculus it is oriented vertically.
Thus no matter what position the head is in, gravity will be acting on one or both sets of otoliths causing deflection of the cell processes on the hair cells and stimulating the sensory nerves at their base. The stimulation is constant due to the ongoing effect of gravity on the mass of the otoliths. Linear acceleration or deceleration will also affect otolith position (Fig. 8.2).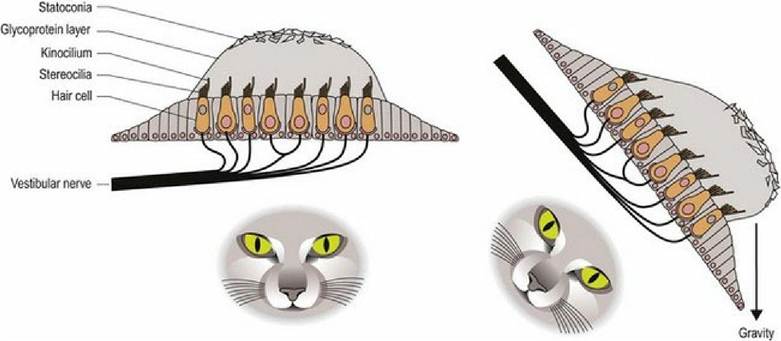
Fig. 8.2 Macula of either the utriculus or sacculus. Regardless of head position gravity will be acting on the mass of the statoconia in one or both maculae. This force pulls on the glycoprotein gel and deflects the
stereocilia towards, or away from, the kinocilium, stimulating or inhibiting, respectively, nerve cells at the base of the hair cells.
Angular acceleration and deceleration of the head
The three semicircular canals and their membranous ducts are located at right angles to each other (in the x, y and z directions) with the anterior and posterior canals being vertically oriented and the horizontal canal being laterally oriented. The semicircular ducts on each side of the head function as synergic pairs. Turning the head in one direction will cause endolymph to flow towards the ampulla on that side, but away from the ampulla in the paired duct on the opposite side. This causes stimulation of hair cells on the first side, but inhibition on the opposite side. Thus the input received by the vestibular nuclei of the brainstem is unequal. Unequal input from paired semicircular canals is interpreted as head movement. Note that if there is disease in the vestibular system on one side, then even at rest the brain will receive uneven input and thus, it is perceived that the animal’s head is moving. The uneven input from the two sides will be interpreted as head movement inducing the reflex changes in posture and eyeball position/movement (see Clinical dysfunction) that result in clinical signs of vestibular disease.
Like the semicircular canal, the semicircular duct is dilated at one end forming the ampulla. The hair cells that perceive head movement are located on a ridge of tissue (crista ampullaris) in the ampulla of the semicircular ducts. The numerous stereocilia and a single kinocilium project from the surface of the hair cells into a gelatinous matrix called the cupula; this projects into the endolymph.
As the head moves, so will the petrous temporal bone, and the associated semicircular ducts and hair cells (Fig. 8.3 and 8.4). However, due to inertia, movement of the endolymph inside the ducts lags behind. The endolymph presses on the cupula, which deflects the stereocilia towards or away from the kinocilium, resulting in stimulation or inhibition of the vestibular neurons, respectively.
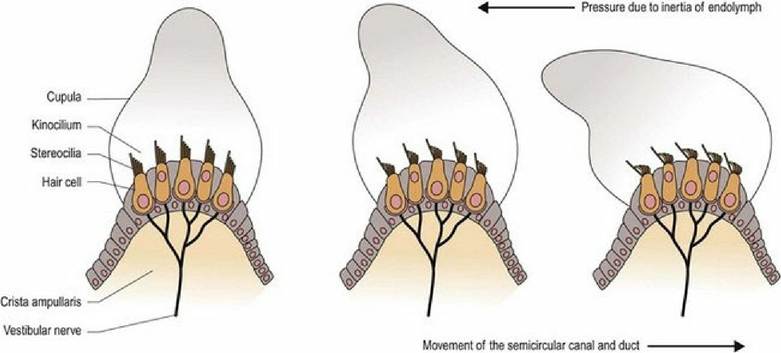
Fig. 8.3 Deflection of processes on hair cells in the ampulla of the semicircular duct, by endolymph pushing on the cupula during head movement.
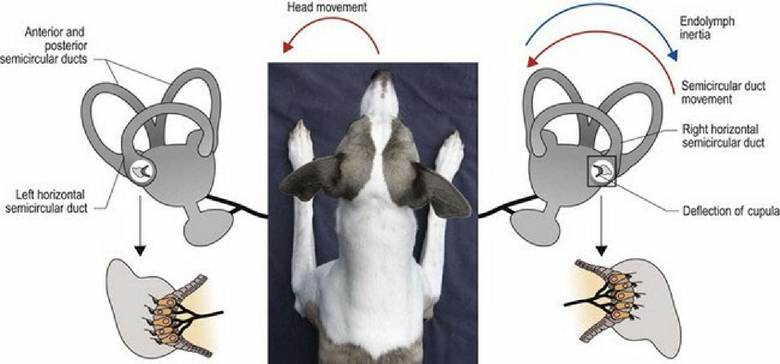
Fig. 8.4 As Timmy turns his head to the left, endolymph in the paired horizontal canals flows in opposite
directions, causing stimulation in the left canal and inhibition in the right canal. The brain receives uneven input from the two canals and this is interpreted as angular movement of head to the left.
If the head continues to rotate in the same direction at the same speed, endolymph movement will catch up with the movement of the ducts and hair cells. As it no longer exerts pressure on the cupula, the stereocilia will not be deflected and the vestibular neurons will not be stimulated or inhibited. Thus there is minimal detection of head movement, via the semicircular ducts, when the head is moving at constant velocity.
When the animal stops rotating, the duct and hair cells cease moving, but again, due to inertia, the endolymph continues to move, but in the opposite direction relative to the membranous labyrinth.
The cupulas in the synergic ducts are deflected in the opposite direction. Again there is uneven input to the vestibular nuclei and this is interpreted as head rotation but in the opposite direction. This causes the sensation of dizziness that humans perceive.Effect of stimulating the vestibular nuclei
The cell bodies of the bipolar, vestibular neurons are located in the vestibular ganglion in the petrous temporal bone. The efferent axons from these neurons join with those from the cochlear portion to form the vestibulocochlear nerve (CN VIII). CN VIII enters the cranial vault through the internal acoustic meatus. It attaches to the lateral aspect of the medulla oblongata just caudal to the caudal cerebellar peduncle.
There are four vestibular nuclei (rostral, medial, lateral and caudal) located near the caudal cerebellar peduncle on each side of the brainstem. Outputs from the vestibular nuclei project to the spinal cord, cerebellum, brainstem and forebrain (Fig. 8.5).
Fig. 8.5
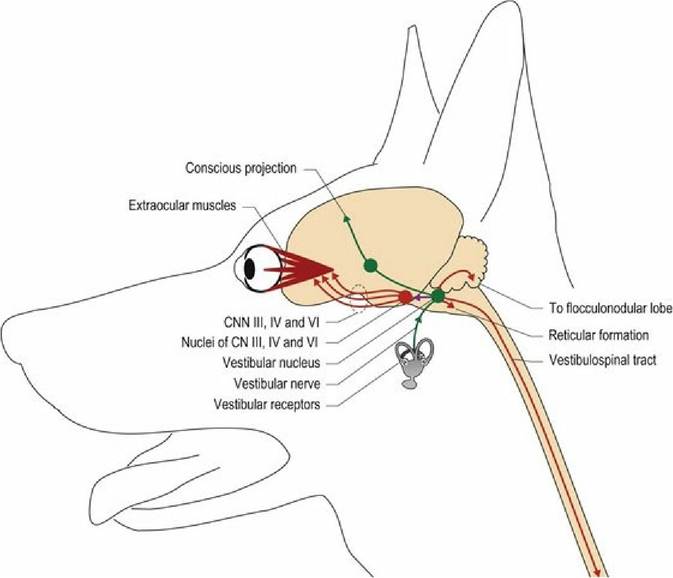
Neural connections arising from the vestibular nuclei of the brainstem.
An animal turning its head to the left illustrates the effects of these outputs.
1. Posture. Vestibular nuclear projections to the spinal cord are via the vestibulospinal tracts and medial longitudinal fasciculus in the ventral funiculus. The fibres terminate on interneurons that facilitate ipsilateral extensor muscles and inhibit ipsilateral flexors. Fibres also decussate to inhibit contralateral extensor muscle activity. Vestibulospinal tracts facilitate spinal reflexes, especially those involved in maintaining the posture (antigravity/extensor muscles). Turning the head to the left causes a shift in the distribution of body mass to the left; this shift in mass is supported by increased extension on the left side. The extension is reflexively induced involving both the myotatic (see Chapter 5) and vestibular reflexes.
Extension on the right side is reduced, thereby minimising the animal’s weight transfer to the left (Fig. 8.6). The vestibulospinal (and reticulospinal) tracts are powerful facilitators of the antigravity/extensor muscles. This influence is demonstrated by the rigidity observed in decerebrate animals in which there is loss of inhibition of the vestibular (and reticular) nuclei.2. Subconscious proprioception. Vestibulocerebellar tracts project to the cerebellum and convey information about head position and movement. The portion of the cerebellum that receives vestibular input is called the vestibulocerebellum and is sited caudoventrally in the flocculonodular lobe (see Chapter 7). The cerebellum uses this information about head proprioception (both static and dynamic), in conjunction with proprioceptive input from the body and limbs, to modify and coordinate motor output from UMN centres of the brainstem and forebrain, to maintain static and dynamic posture of the whole body.
3. Eyeball position and movement. Fibres from the vestibular nuclei project rostrally (in the medial longitudinal fasciculus/vestibulomesencephalic tract) to synapse on cranial nerve nuclei III, IV and VI that supply LMN to the extraocular muscles (Chapter 10). This wiring is the basis of the vestibulo-ocular reflex in which head movement cause reflex changes in eyeball position, and sets the position of the globes in the orbits at the end of head movement. During head movement the eyeballs move reflexively in a jerky fashion, called physiological nystagmus. The jerky movement is due to fixation of the gaze on a spot for a moment before flicking rapidly to a new position. Physiological nystagmus enables the animal to maintain a useful visual input during head movement. If the eyeballs remained centred in the orbit during movement, then the visual field would become blurred. For example, an animal is gazing at a spot in front of it, but then begins to move its head to the left. Its gaze remains fixated on that first spot, thus both eyes deviate to the right as the head moves to the left.
The left lateral rectus muscle and right medial rectus muscle of the eyes are increasingly stretched until suddenly, and reflexively, they are stimulated to contract; this moves the eyes abruptly to the left. The gaze is fixated on a new position, the head continues to turn, muscles are stretched and rapid eyeball movement to the left again occurs, etc. Thus the animal’s eyes deviate slowly in the opposite direction to which the head is turning and then make a rapid, active movement in the direction that the head is turning (see Fig. 13.7). This rhythmical, reflexive eyeball movement is called physiological nystagmus. Similarly, if the animal moves its head up or down, vertical nystagmus is induced with the fast phase of eyeball movement in the direction that the head is moving. The input from the vestibular system and output to CNN nuclei III, IV and VI also set the position of the eyes at the end of head movement. The position of the globe will be centred in the orbit after horizontal movement, but after vertical movement the default position will depend on whether it is a predator or prey species. In carnivores, the eyes will be centred in the orbit, thus if the head is tilted upwards, the eyes will also be looking upwards. In herbivores, with an elevated head position, the eyes will be directed downwards as the default normal position. Professor R. Le Couteur, UC Davis, teaches a useful way of remembering this. As the big cat is chasing the herbivore in the wild and is going to leap for the throat, the cat has its head tilted upwards and is looking forward (eyeballs centred in orbit) at that luscious target of the throat. The herbivore has its head tilted up as far out of reach as possible, but is looking down at the slavering jaws of the predator snapping below it.4. Conscious awareness. Other rostrally directed fibres travel bilaterally, synapsing in the medial geniculate nucleus of the diencephalon, and then via the internal capsule to the somatosensory cortex for conscious awareness of head position.
5. Motion sickness. Local projections from the vestibular nuclei into the brainstem connect to the vomiting centre of the reticular formation. This is the pathway by which head movement may trigger motion sickness, which occurs when visual and vestibular inputs do not correlate with each other.

Fig. 8.6 As Barney turns to the left, uneven stimulation of the vestibular nuclei results in ipsilateral extension, inhibition of ipsilateral flexion and contralateral extension. Thus a change in posture supports the shift in the centre of gravity.
Note that there is also input to the vestibular nuclei from the first three cervical segments of the spinal cord utilising the spinovestibular tract in the ventral funiculus. The input conveys proprioceptive information about head position from muscles at the head-neck junction to the caudal vestibular nucleus. Thus, damage to the first three cervical vertebral spinal cord segments, or their dorsal roots, can also result in dramatic vestibular signs.
Also note that in the cat and other types of mammal, up to 20% of fibres in the vestibular nerve have been described as efferent, originating from a variety of regions in the medulla oblongata, including the reticular formation and the vestibular nuclei. Their role may include facilitation, or inhibition, of vestibular receptors in the membranous labyrinth.
Relationship between the vestibular apparatus and neck reflexes
There is an intimate relationship between head position, neck position and limb position. If head position alters, without changing the head-neck angle, the primary input is from the vestibular apparatus, which induces reflex activity in the limbs and trunk; these are called vestibular reflexes. Changes in head-neck junction angles are detected by proprioceptors in the neck muscles. The resulting reflex changes in the limbs are induced by neck reflexes.
Neck muscles at the head-neck junction, such as the rectus and obliquus capitus muscles, are rich in muscle spindles with five times the number of muscle spindles per gram compared with a large, postural muscle like the gluteals. Input from these and other neck muscles, travel cranially in the spinal cord via the spinovestibular tract to the caudal vestibular nuclei. Spinovestibular fibres are intermingled with the lateral vestibulospinal tract fibres in the ventral funiculus. The overall effect of vestibular and neck reflexes on limb flexion and extension is summarised in Fig. 8.7.
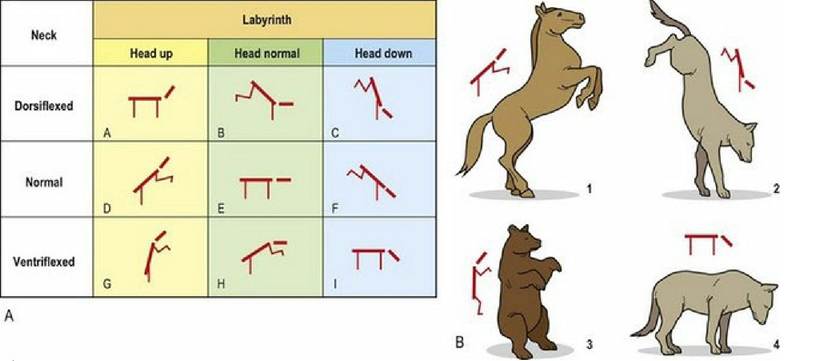
Fig. 8.7 Limb position is reflexively induced by sensory input from both the vestibular apparatus (labyrinth) and from proprioceptors in the muscles at the head neck junction. Hence an animal’s limb posture will be dependent on whether the head is tilted or neutral, and whether there is flexion or extension at the head-neck junction.
(Figure redrawn with permission from Roberts TDM, Understanding Balance, The Mechanics of Posture and Locomotion. Chapman & Hall, London, 1995).