Synaptic Transmission
Synapses are specialized junctions where information is exchanged between neurons or between a neuron and the cell or cells that it innervates. Two general types of synapses, electrical and chemical, are found between neurons in the nervous system, with chemical synapses being more prevalent.
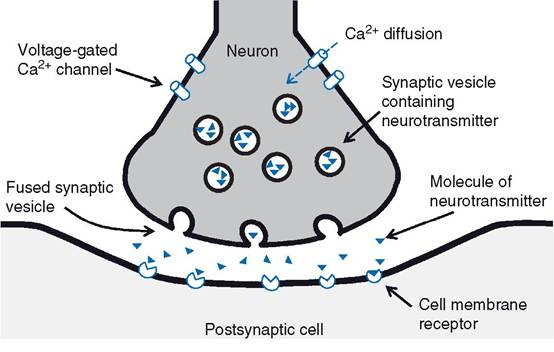
Electrical synapses are essentially gap junctions between the cell membranes of adjacent neurons that permit ionic exchange.information exchange at chemical synapses entails the release of a chemical neurotransmitter from the presynaptic neuron (Fig. 10-3). When an action potential arrives at the terminal end of the presynaptic neuron, the change in membrane potential is believed to be responsible for opening voltage-gated calcium channels. The calcium concentration within the cytosol is lower than the calcium concentration in the extracellular fluid, so calcium can diffuse through the open channels into the cell down its concentration gradient. The increase in intracellular calcium within the terminal end of the presynaptic neuron is associated with exo- cytosis of neurotransmitters stored in secretory vesicles within the presynaptic neuron. Typically, an individual neuron contains vesicles with only one primary neurotransmitter, but a wide variety of substances have been found to function as neurotransmitters. These are discussed later in this chapter.
The cell membranes of presynaptic and postsynaptic neurons (or other target cells) are not in immediate contact. A small but distinct separation exists, and the space between is the synaptic cleft. Neurotransmitters released by the presynaptic neuron must diffuse across the synaptic cleft to have their effect on postsynap- tic neurons, or target cells. However, this diffusion occurs almost instantaneously because of the very small size of the cleft (average 20 nm, or about a millionth of an inch).
The events of synaptic transmission are summarized in Figure 10-3.Neurotransmitters typically bind to cell membrane receptors on postsynaptic neurons or target organs. At synapses between neurons, the binding changes the postsynaptic membrane’s permeability to ions (either directly or indirectly via second messengers), and this in turn produces a change in the membrane potential of the postsynaptic neuron. synapses and the neurotransmitters that depolarize the postsynaptic neuron are excitatory synapses and excitatory neurotransmitters, while synapses and neurotransmitters that hyperpolarize the postsynaptic neuron are inhibitory. The change in postsynaptic membrane potential produced by excitatory neurotransmitters is an excitatory postsynaptic potential (EPSP), while an inhibitory neurotransmitter produces an inhibitory postsynaptic potentials (IPSP). Typically, excitatory neurotransmitters cause membrane depolarization by increasing the membrane’s permeability to sodium, while inhibitory neurotransmitters increase the membrane’s permeability to either potassium or chlorine. Hyperpolarization of the cell membrane can be brought about by either an increased rate of potassium exit from a cell or an increased rate of chlorine entry into the cell down their respective concentration gradients.
Figure 10-3. Summary of events involved in synaptic transmission. Action potential arrives and promotes the opening of voltage-gated calcium channels. Entry of calcium promotes fusion of synaptic vesicles that release neurotransmitter. Neurotransmitter diffuses to postsynaptic cell membrane and binds to its cell membrane receptor.
The amount of excitatory neurotransmitter released by a single action potential in a single presynaptic neuron is constant. However, it is also typically insufficient to depolarize the postsynaptic neuron to a threshold voltage at which an action potential can be elicited.
Therefore, to reach threshold voltage in a post- synaptic neuron, the change in voltage produced by multiple EPSPs must be summated. Summation of EPSPs is classified as either temporal or spatial. Temporal summation occurs when a single presynaptic neuron releases neurotransmitter repeatedly before the effect of each single release is lost. When effective summation occurs, the additive effects of the multiple releases are enough to produce an action potential in the postsynaptic neuron. Spatial summation occurs when simultaneous or nearly simultaneous neurotransmitter release occurs at more than one synapse on the postsynaptic neuron. Spatial summation is possible because neuron cell bodies typically have multiple junctions with many presynaptic neurons.After a neurotransmitter has been released and had its desired effect on the postsynaptic neuron, it must be removed to prevent continuous stimulation of the postsynaptic neuron. The specific mechanisms by which neurotransmitters are removed from neural synapses vary among neurotransmitters. However, in general these can be any one or a combination of the following: (1) enzymes in the area of the synapse degrade the neurotransmitter; (2) cell membrane transport systems absorb the neurotransmitter; or (3) the neurotransmitter diffuses away from the area of the synapse.
Typically, as described earlier, a single neuron and its dendrites contain multiple synaptic junctions and receive synaptic input from multiple presynaptic neurons, so a single neuron may receive impulses from several sources. This pattern or organization is described as convergence (Fig. 10-4). Divergence is the opposite: each axon branches so that synaptic connections are made with many neurons (Fig. 10-4). These organizational patterns permit information to be widely distributed throughout a neural network (divergence) or permit multiple sources of information to be brought to focus on a single neuron for a focused response (convergence). These are very simple illustrations of how the organization of neuronal networks can contribute to the processing and integration of information. For example, a single neuron can be simultaneously stimulated by excitatory and inhibitory neurotransmitters from different presynaptic neurons (a converging network). The property of spatial summation permits the neuron receiving these converging inputs to integrate the different stimuli and respond appropriately.
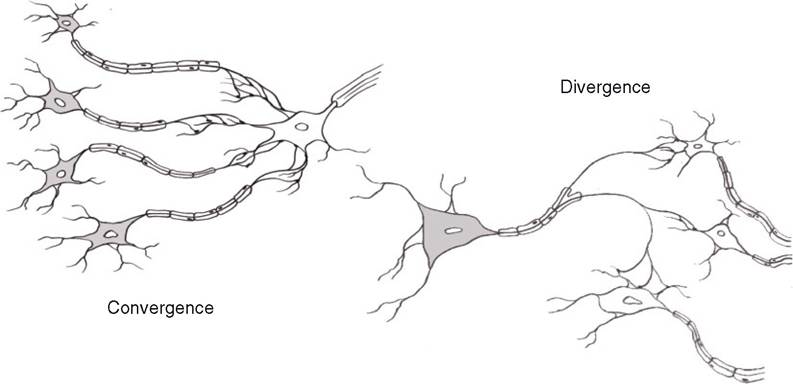
Figure 10-4. Simple patterns to illustrate convergence and divergence in neural networks.