THE CONTENTS OF THE VISCERAL SPACE
The visceral space of the neck is enclosed by four superficial and two deep muscles. The sternohyoid muscle ventral to the trachea extends from the manubrium to the basihyoid; it is loosely connected with its fellow in the midline.
The sternothyroideus, also thin and straplike, lies lateral to the trachea, ending on the lateral surface of the thyroid cartilage. These are the only structures that intervene between the larynx and trachea and the skin in the cranial half of the neck (Figure 11-45). They are covered by the sternocephalicus in the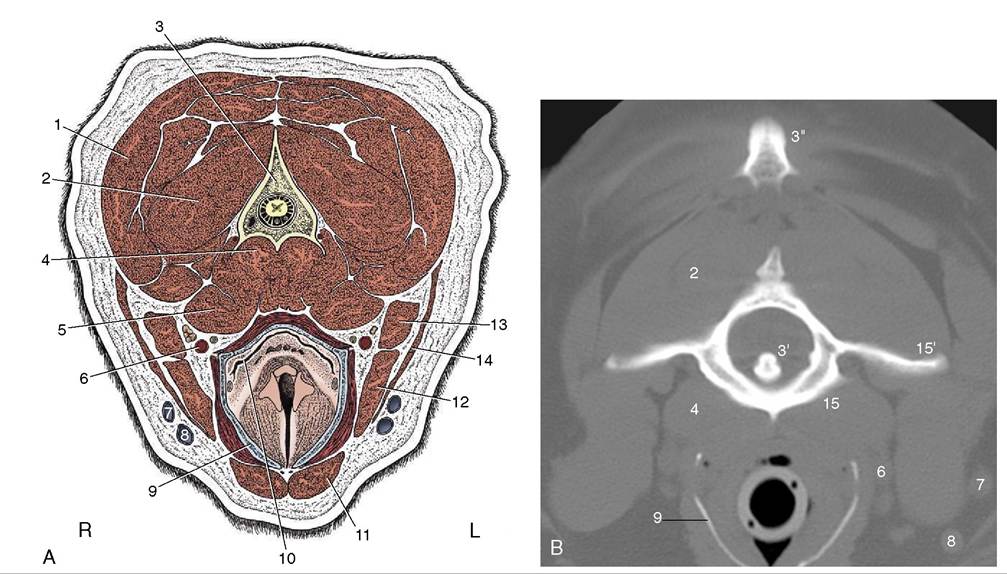
Figure 11-45 A, Transverse section of the canine neck at the level of the axis. B, Corresponding CT scan (bone window), slightly more cranial than A. 1, Splenius; 2, obliquus capitis caudalis; 3, axis; 3', dens of axis; 3", cranial tip of spine of axis; 4, longus colli; 5, longus capitis; 6, common carotid artery, vagosympathetic trunk, and medial retropharyngeal lymph node; 7, maxillary vein; 8, linguofacial vein; 9, thyroid cartilage (calcified); 10, laryngopharynx, leading into esophagus; 11, sternohyoideus; 12, sternomastoideus; 13, cleidomastoideus; 14, sternooccipitalis; 15, atlas; 15', wing of atlas.
caudal half. This muscle consists of two parts, the sternomastoideus and the sternooccipitalis, which diverge toward the head (Figure 11-46). The dorsal sternooccipitalis ends on the back of the skull.
The brachiocephalicus also has two parts in the neck, the cleidomastoideus and the cleidocervicalis. The former passes deep to the sternooccipitalis to a common insertion with the sternomastoideus on the mastoid process of the temporal bone. The latter sweeps over the lateral surface of the neck to meet its fellow in the dorsal midline (see Figure 2-55/2).
The sternocephalicus and brachiocephalicus are fused except caudally, where there separation allows the external jugular vein to become more superficial (see Figure 11-46).
The deep muscles comprise the longus capitis, ventrolateral to the cervical vertebrae, and the longus colli more medially (Figure 11-45/4,5). The fascia that covers these muscles ventrally detaches a superficial leaf that encloses the many structures in the visceral space: the esophagus, trachea, thyroid and parathyroid glands, common carotid arteries, vagosympathetic trunks, internal jugular veins, recurrent nerves and tracheal lymph nodes (see Figure 11-46, A). There is no cervical component of the thymus.
The esophagus continues from the laryngopharynx. It first lies dorsal to the trachea, but it deviates to the left in the middle of the neck and maintains this position through the thoracic inlet. The esophagus and trachea are thus both in contact with the longus colli in the caudal half of the neck. The esophagus may be felt with the fingertips as a pliable tube sinistrodorsal to the trachea. The habit of dogs of bolting their food often leads to obstructions. Large pieces of meat, gristle, or bone—and not infrequently stones—tend to lodge at the thoracic inlet, where the esophagus is unable to expand fully.
The trachea continues from the larynx and, because of its firmness, is easily palpated. Unlike the esophagus, it can be grasped so that the flat dorsal surface between the ends of the tracheal rings can be appreciated. In normal dogs it may be possible to demonstrate modest changes in tracheal diameter in synchrony with the phases of respiration. The cervical trachea, especially its caudal part, narrows slightly during inspiration only to recover during expiration. The changes in the tho-
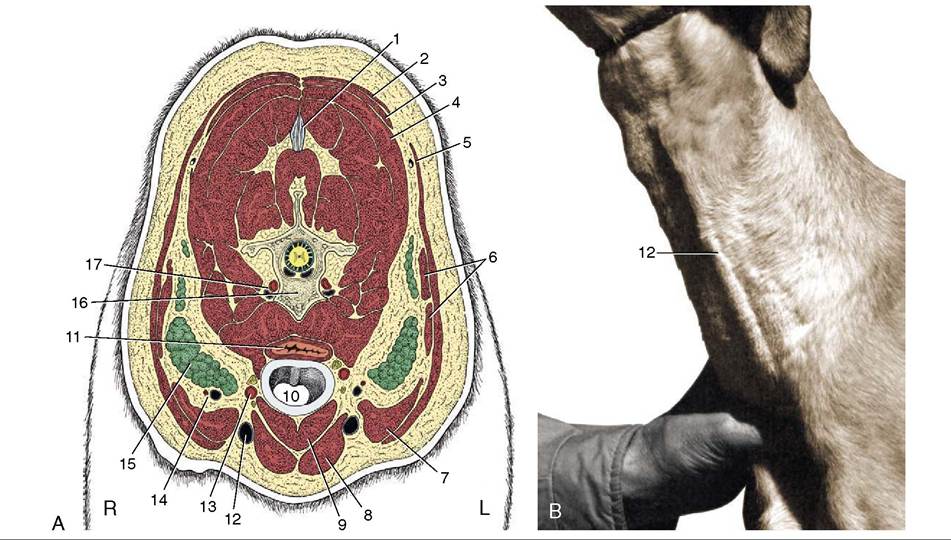
Figure 11-46 A, Transverse section of the canine neck at the level of the fifth cervical vertebra. B, Left external jugular vein raised by thumb pressure at the base of the neck. 1, Nuchal ligament; 2, trapezius; 3, rhomboideus; 4, splenius; 5, cleidocervicalis; 6, omotransversarius; 7, cleidomastoideus; 8, sternocephalicus; 9, sternothyrohyoideus; 10, trachea; 11, esophagus; 12, external jugular vein; 13, common carotid artery, vagosympathetic trunk, and recurrent laryngeal nerve; 14, superficial cervical vessels; 15, superficial cervical lymph nodes; 16, fifth cervical vertebra; 17, vertebral vessels.
racic trachea are reciprocal. This physiological variation is not to be confused with the more severe narrowing of the tracheal lumen, possibly amounting to collapse, that sometimes develops with congenital or acquired degeneration of the supporting cartilages. In this pathological condition the cervicothoracic transitional portion of the trachea is most often affected.
There is radiographic evidence that brachycephalic breeds have relatively narrow tracheas, whereas Dachshunds and Basset Hounds have wide ones. Estimates of the normality of the tracheal diameter may be made by comparing it with the height of the thoracic inlet; in some breeds the ratio may be as high as 0.5, whereas in severely affected Bulldogs it may be as low as 0.05.
The trachea is loosely enclosed in a sleeve of fascia. A deeper leaf forms part of the prevertebral fascia that separates the trachea from the longus colli muscle. It also contributes to the carotid sheath, which encloses the vagus and sympathetic nerves, carotid artery, internal jugular vein, and sometimes the tracheal lymph trunk. The carotid sheath is found dorsolateral to the trachea; the recurrent laryngeal nerve follows a similar but independent course.
Each tracheal ring is thickest ventrally and thins along the curves to end dorsally as flexible, potentially overlapping blades. Only the first ring is completely closed in dogs and is partially covered by the cricoid cartilage. The dorsal part of the trachea is composed of connective tissue and muscle. In carnivores, this smooth muscle inserts on the external surface of the cartilages some distance from their tips.
The thyroid gland consists of two elongated rather flattened lobes placed against and loosely attached to the lateral aspects of the first few tracheal cartilages under cover of the sternothyroid muscle (see Figure 6-4, A). Their caudal poles are sometimes connected across the ventral surface of the trachea by a vestigial isthmus. They are embedded in the deep cervical fascia.
The sternocephalicus and sternohyoideus muscles pass immediately lateral to the convex surface of each gland, and the sternothyroideus covers each thyroid ventrally. The recurrent laryngeal nerve passes dorsally. In medium-sized dogs the lobes are about 5 cm long (spanning the first five to eight tracheal rings) and 1.5 cm wide. In immature dogs and in those of brachycephalic breeds, they are larger. In cats each thyroid gland lobe is about 2 cm long and 0.3 cm wide. Frequently, accessory thyroid tissue occurs along the trachea at the thoracic inlet, within the mediastinum, and along the thoracic portion of the descending aorta. During devel-opment islets of the rapidly proliferating cells of the thyroid primordium separate from the main mass and become incorporated in the developing structures of the branchial arch region and thorax.
The major blood supply to each lobe is provided by a cranial thyroid artery (branching from the common carotid artery), a vessel with a larger distribution than its name suggests. Its thyroid branches include one that follows the dorsal margin caudally to an anastomosis with the much smaller and inconsistent caudal thyroid artery (branching from the brachiocephalic artery); one that follows the ventral margin; and others that pass directly to the cranial pole (and to the external parotid gland). Twigs from all these result in the thyroid being supplied at scattered points around most of its periphery. Blood leaving the gland enters the nearby internal jugular vein, while some is conveyed to the large veins at the thoracic inlet by an unpaired (caudal thyroid) vein lying on the ventral surface of the trachea.
Each lobe is closely associated with two parathyroid glands (discounting the possible existence of accessory parathyroid tissue) in a relationship of obvious relevance to the performance of thyroid surgery. The external (III) parathyroid gland is generally found close to or against the cranial pole of the thyroid to which it is loosely joined; in cats more often than in dogs, this glands descends unusually far from its site of origin (p.
220) and comes to rest near the caudal pole. The internal (IV) parathyroid is located within the connective tissue capsule of the thyroid and may be difficult to discover, especially when completely submerged within thyroid glandular tissue, as sometimes happens. Recognition is assisted by its pale color, which contrasts with the brownish-red thyroid tissue; it can be identified on or within the thyroid gland by ultrasonography. The sizes of the parathyroid glands are rather variable, but on average they are about 3 mm in diameter in dogs. Partial or complete thyroidectomy may be performed in the treatment of thyroid hyperplasia or neoplasia, the former condition now recognized as occurring with great frequency in cats. Certain surgical procedures (intracapsular thyroidectomy) make the concomitant removal of a considerable fraction of the total parathyroid tissue more or less inevitable; this loss is generally tolerated, provided the integrity of the blood supply to the remaining part is preserved. Caution is obviously most necessary when surgery is bilateral.The common carotid artery runs dorsolateral to the trachea (though the left one is commonly displaced to the side of the esophagus in the caudal half of the neck). It arises from the brachiocephalic trunk about 1 cm apart (sometimes a bicarotid trunk is formed) within the chest and crosses the lateral surface of the trachea (esophagus on the left) obliquely to gain a dorsolateral position in the neck. The cranial thyroid artery, which arises level with the larynx, is the only cervical branch of consequence. The common artery ends at the level of the atlantooccipital joint by dividing into internal and external carotid arteries. The former enters the skull through the carotid foramen after pursuing a rather unusual course (p. 311).
The internal carotid artery (much smaller than the external one) leaves the medial side of the parent vessel and almost at once displays the bulbous enlargement, the carotid sinus (see Figure 7-32).
It makes its way between deep structures of the head, crossing the lateral surface of the pharynx, without detaching any branches and enters rostral to the tympanic bulla in the skull to supply the brain. The internal carotid artery is regressed in the adult cat; the main blood supply to the brain is then coming from branches of the maxillary artery.The external carotid artery forms a sigmoid flexure as it winds its way under the hypoglossal nerve, submandibular salivary gland, and the digastric muscle. Its many branches include the occipital, cranial laryngeal, ascending pharyngeal, lingual, facial, caudal auricular, parotid, superficial temporal, and maxillary arteries (see Figure 7-39). The occipital sometimes arises independently from the common carotid.
The internal jugular vein is formed by the confluence of the vertebral vein, the sigmoid sinus, and, occasionally, the vein of the hypoglossal canal. The internal jugular is first associated with the internal carotid artery in the sheath of the common carotid. This vein usually terminates in the caudal part of the external jugular vein, which is the main channel for venous return from the head. It begins by the union of the linguofacial and maxillary veins. In the adult it contains a few ineffective, irregularly spaced valves.