The Development of the Heart
The primitive heart, the single median structure formed by the fusion of paired rudiments, is carried ventral to the foregut by the reversal process reshaping the head end of the embryo (p.
91). Though initially consisting of a simple endothelial tube, the heart soon acquires an investment of mesoderm that forms the myocardial and epicardial components of its wall. The cranial part of the tube, which will later form the truncus arteriosus and ventricles, is at this stage contained within the pericardial cavity and suspended by a fold (dorsal mesocardium) extending between the myoepicardium and the pericardial wall (Fig. 7.1B/9). The caudal part, which forms the atria and sinus venosus, first lies caudal to the pericardial cavity embedded within the septum transversum. The enclosed (truncoventricular) part of the heart grows more rapidly than the pericardial space and is forced into a flexure whose apex is directed ventrocaudally and somewhat to the right. The atrial expansions of the initially paired endothelial tubes have now fused in a single common atrium continuous with the sinus venosus; this presents an unpaired transverse part that receives the paired horns created by the entry of the veins (Fig. 7.25).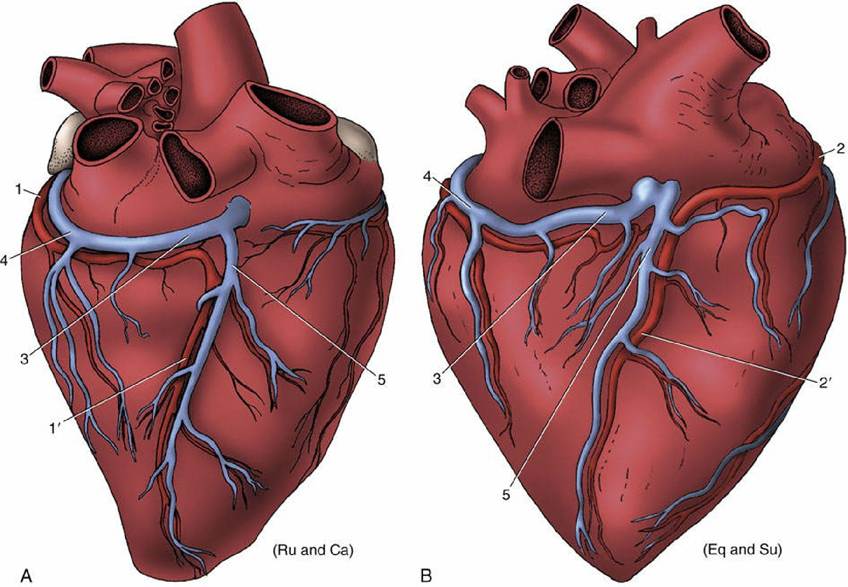
FIG. 7.20 Patterns of coronary circulation of the heart viewed from the right. (A) Situation in ruminants (Ru) and carnivores (Ca); the right (subsinuosal) interventricular branch (1') is a continuation of the left coronary artery. (B) Situation in the horse (Eq) and pig (Su); the right (subsinuosal) interventricular branch (2) is a continuation of the right coronary artery. 1, Circumflex branch of left coronary artery; 2, right coronary artery; 3, coronary sinus; 4, great cardiac vein; 5, middle cardiac vein.
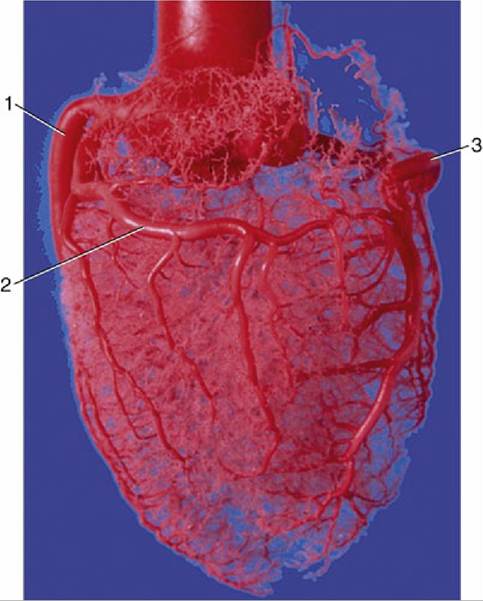
FIG.
7.21 Corrosion cast of aorta and coronary circulation (pig). 1, Left coronary artery; 2, ramus circumflexus; 3, right coronary artery.
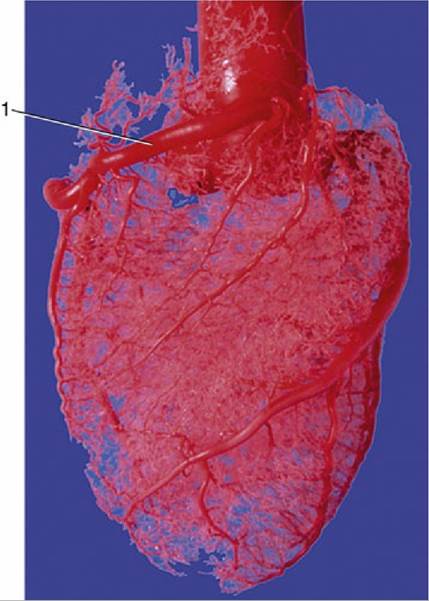
FIG. 7.22 Corrosion cast of aorta and coronary circulation (pig). 1, Right coronary artery.
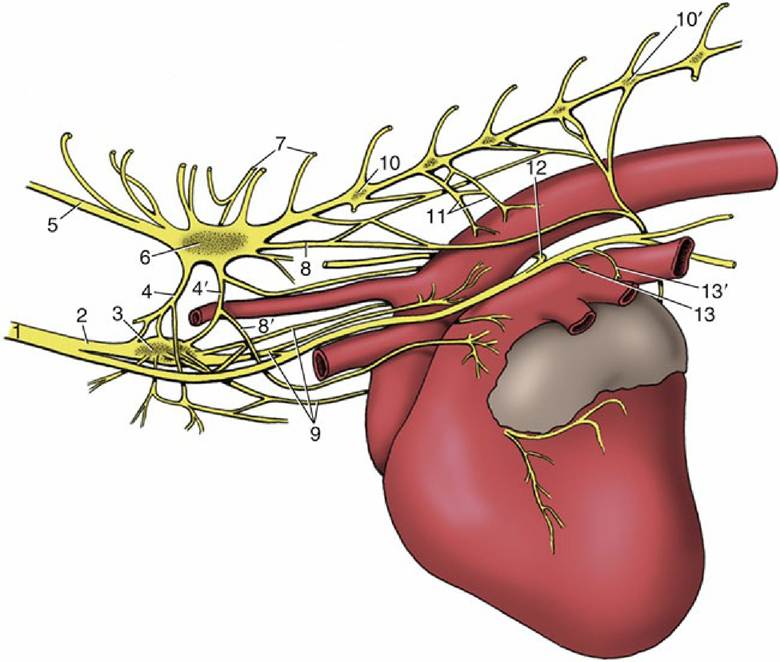
FIG. 7.23 Cardiac nerves and related ganglia of the dog; left lateral view. 1, Vagosympathetic trunk; 2, sympathetic trunk; 3, middle cervical ganglion; 4 and 4', cranial and caudal limbs of ansa subclavia; 5, vertebral node; 6, cervicothoracic ganglion; 7, communicating branches; 8 and 8', caudodorsal and caudoventral cervicothoracic cardiac nodes; 9, vertebral cardiac nodes; 10 and 10', third and seventh thoracic ganglia; 11, thoracic cardiac nodes; 12, left recurrent laryngeal node; 13 and 13', cranial and caudal vagal cardiac nodes.
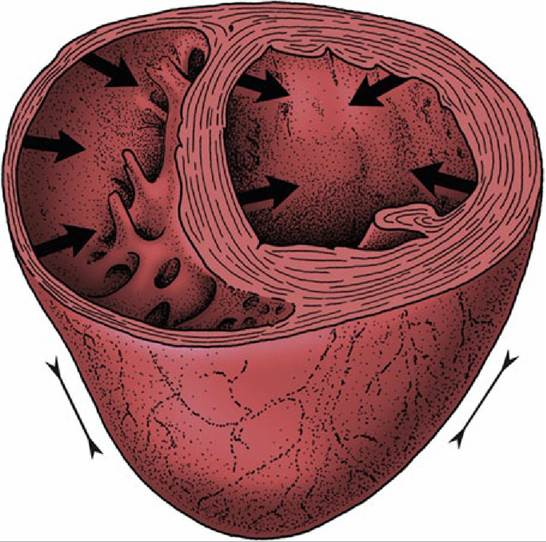
FIG. 7.24 Schematic drawing of the mode of contraction of the left and right ventricles. The wall of the left ventricle contracts radially, whereas the right ventricular lumen is squeezed in a bellows action. Arrows indicate direction of contraction.
The four heart chambers at this stage, in caudocranial sequence, are sinus venosus, atrium, ventricle, and truncus arteriosus. The atrium and ventricle are separated by a constriction called the atrioventricular canal, whereas the transition from ventricle to truncus forms the arterial conus (bulb of the heart). The truncus continues rostrally into the aortic arches, which now appear in the mesoderm to each side of the pharynx (see Fig. 7.2B). The sinus venosus receives the cardinal, vitelline, and umbilical systems of veins that extend from the body of the embryo, the yolk sac, and the chorioallantois, respectively (see Fig. 7.25). The bifid character of the sinus venosus persists for a time, but its wide communication with the atrium gradually shifts toward the right as the amount of blood entering the left horn is diminished after the obliteration of the left umbilical and left vitelline veins.
When the sinus is eventually incorporated within the atrium, the undivided part and the right horn contribute the sinus venarum of the adult right atrium, but the left horn is reduced to the coronary sinus. By this stage the sinus venosus and common atrium have also become included within the pericardial cavity, where they lie dorsal to the ventricle.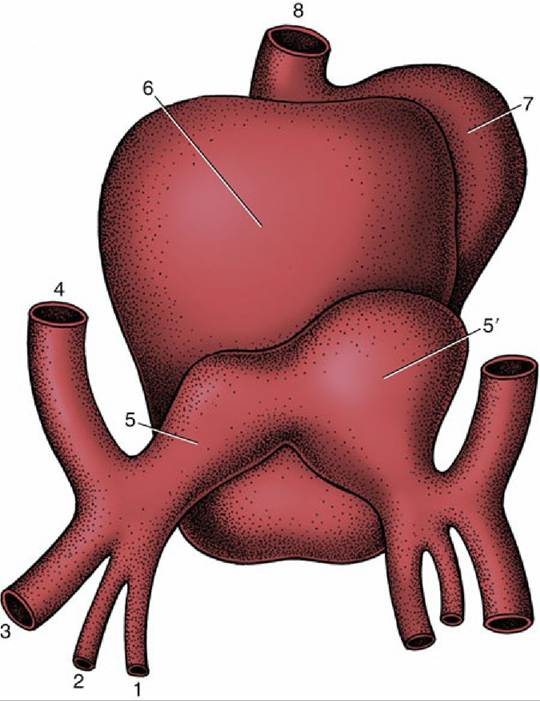
FIG. 7.25 Dorsal view of the developing heart. 1, Vitelline vein (v.); 2, umbilical v.; 3, caudal cardinal v.; 4, cranial cardinal v.; 5 and 5', left and right horns of sinus venosus; 6, atrium, 7, ventricle; 8, truncus arteriosus.
The appearance and subsequent growth of a crescentic ridge, known as the septum primum, initiates the division of the common atrium into right and left chambers (Fig. 7.26/2). This ridge projects ventrally into the lumen and at its ends grows toward thickenings of the wall of the atrioventricular canal known as the endocardial cushions (Fig. 7.26/6). The opening between its free margin and the cushions is known as the ostium primum (Fig. 7.26/4). The ostium primum is gradually occluded by the further enlargement of the cushions, but before closure is complete, a number of perforations appear within the septum and coalesce to form a fresh communication, the ostium secundum (Fig. 7.26/5), between the two atria. The definitive division of the atria is achieved by a second crest (Fig. 7.26/3) that now appears to the right of the primary partition. The concave free ventral margin of this second crest overlaps the ostium secundum, reducing the passage between the atria to a narrow space, the foramen ovale, between the two septa (Fig. 7.26C). The remnant of the septum primum forms the valve of the foramen ovale. The final closure of the opening is accomplished after birth by the apposition and subsequent fusion of the valve to the septum secundum (p. 241).
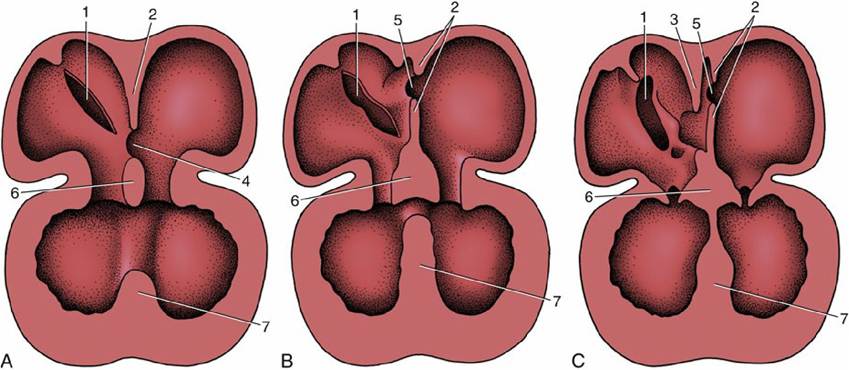
FIG.
7.26 (A) to (C) The partitioning of the atrium and ventricle, schematic. (A) The primary atrial septum has formed, and development of the interventricular septum has begun. (B) The primary atrial septum has fused with the endocardial cushions, and a secondary foramen (5) has been formed. (C) The secondary atrial septum has formed, and a passage (foramen ovale) between primary and secondary septa connects the right and left atria. Note the fusion of the interventricular septum with the endocardial cushions. 1, Sinoatrial opening; 2, primary atrial septum; 3, secondary atrial septum; 4, ostium primum; 5, ostium secundum; 6, fused endocardial cushions; 7, interventricular septum.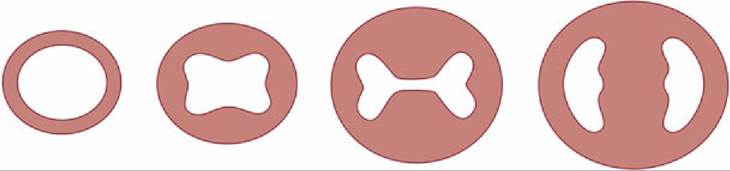
FIG. 7.27 Left to right, Partitioning of the atrioventricular canal by the endocardial cushions. The single atrioventricular canal is gradually divided into right and left atrioventricular openings.
Further growth and eventual mergence of the endocardial cushions divide the canal into the two openings that become the right and left atrioventricular ostia (Figs. 7.27 and 7.28B).
The septation of the truncus arteriosus and bulbus is achieved by the appearance, growth, and fusion of two endocardial ridges that run along the length of the truncus. The left one is known as the septal ridge, the right one as the parietal or dorsal ridge. Fusion of the ridges commences at the distal extremity of the truncus and gradually extends proximally, producing a partition that ends in a free edge arched over the common ventricle (Fig. 7.28B∕2 and 3). The lower end of the parietal ridge expands within the ventricle and contributes to the closure of the atrioventricular ostium. The septal ridge fuses with the most cranial part of the interventricular septum that has been developing in the meantime.
This interventricular septum first appears as a falciform crest formed through local thickening of the myocardium at the apex of the ventricle; as it extends it divides the common cavity into right and left chambers (Fig.
7.26/7). Although the external conformation of the heart at this stage already approximates its final form, the truncus arteriosus (although now divided internally) appears to arise solely from the right ventricle (Fig. 7.28A). The two ventricles still communicate with each other over the free edge of the interventricular septum but are in separate communication with the atria through the paired slitlike openings created by the subdivision of the atrioventricular canal. The right atrioventricular opening is substantially bounded by the right part of the caudal endocardial cushion, less extensively by the cranial cushion, and partly, as already mentioned, by the parietal ridge of the truncus. These three contributions each form a separate cusp of the valve, and the truncus ridge contributes the parietal cusp.The left atrioventricular valve has a similar origin, mainly from the cranial and caudal endocardial cushions but with a small additional (lateral) cushion forming the parietal cusp. The division of the ventricles is largely completed by fusion of the interventricular septum with the caudal cushion; it is finally achieved by fusion of the lower edge of the truncus septum with the right part of the caudal cushion and with the interventricular septum. Because the same process completes the aortic part of the truncus, the output of the heart is now divided into two streams: one from the left ventricle into the aorta and one from the right ventricle into the pulmonary trunk.
Because the process of heart development requires highly precise meeting and fusion of various elements, heart malformations are among the most common congenital abnormalities, affecting up to 1% of all human births. Heart malformations are also frequent in domestic animals. The more common malformations are defects of the cardiac septa, atresia or stenosis of the pulmonary or aortic trunks, or some combination of these anomalies (e.g., the tetralogy of Fallot: pulmonary stenosis, enlarged overriding aorta, ventricular septal defect, and hypertrophy of the right ventricle). Failure of closure of the oval foramen is generally without functional significance, but most other malformations are incompatible with normal life after birth. Surgical correction is neither practicable nor advisable in those affected animals that do not die spontaneously.