The Kidneys Generate New Bicarbonate and Excrete Ammonium Ion
Renal generation and excretion of ammonium ion (NH4+) is a major component in the maintenance of acid-base balance; renal NH4+ excretion is illustrated in Figure 44-2.
In proximal tubule cells the amino acid glutamine is metabolized to produce NH44. This process is called ammoniagenesis. The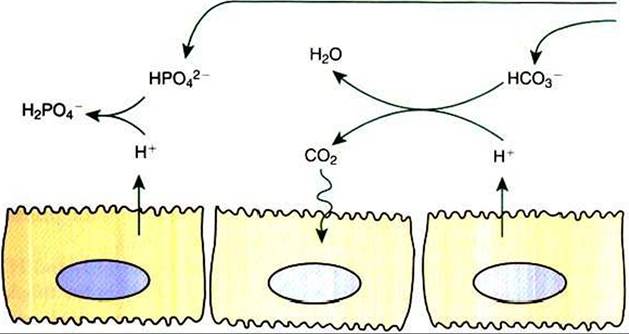
FIGURE 44-1 Schematic illustration of buffer mechanisms at work in tubule fluid. In the proximal tubule, buffering by filtered bicarbonate (HCO3 ) predominates because of the relatively high concentration of HCO3. In the cortical collecting duct, buffering by filtered, nonbicarbonate buffers, such as HPO42, predominates.
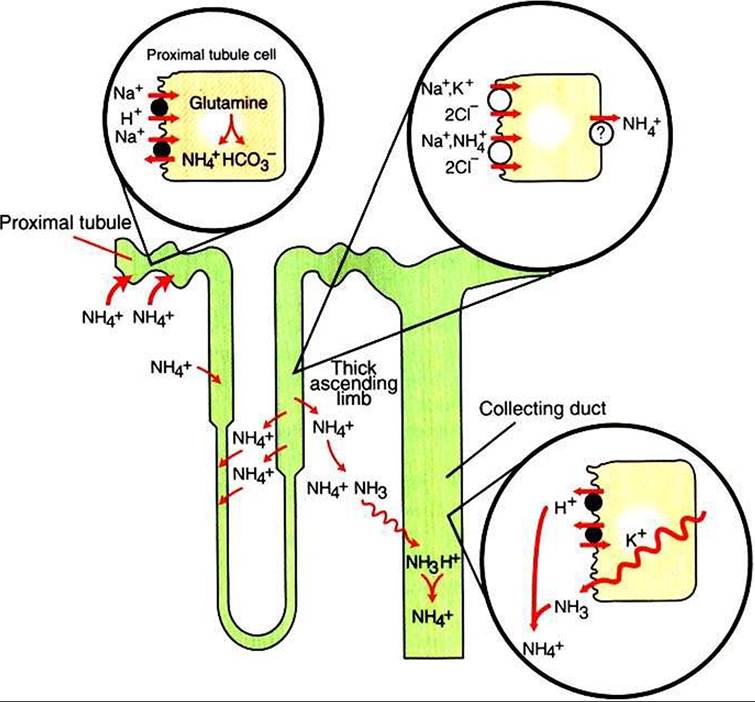
FIGURE 44-2 Schematic illustration of the roles of various nephron segments in ammonium excretion. In the proximal tubule, glutamine is Catabolized to generate ammonium ion (NH4*) and bicarbonate (HCO3 ). NH4* is secreted into the lumen by substitution for H* on the Na*∕H* exchanger in the apical plasma membrane. Ammonium ion recycles in the loop of Henle by reabsorption by the thick ascending limb, in which NH4* is reabsorbed by substitution for K* on the Na*∕K*,2CΓ co-transporter in the apical plasma membrane, which is followed by some form of facilitated transport across the basolateral plasma membrane.The elevation of the interstitial NH4* concentration results in movement into the descending thin limbs of Henle's loop and subsequent return to the thick ascending limb.This medullary recycling results in a high concentration of ammonia (NH3) and NH4* in the medullary interstitium and prevents its return to the cortex, where it would be reabsorbed into the blood.
Ammonia readily diffuses into the collecting duct, where it is rapidly protonated, trapped in the lumen, and then excreted in the urine.intracellular NH4+ enters the tubule fluid through secondary active transport by substitution for H' on the Na+∕H+ exchanger. Glutamine metabolism also produces new bicarbonate anions. Thus the net result of the renal generation and excretion of NH4* is acid excretion and bicarbonate production. Renal ammoniagenesis is enhanced by acidosis and is an important renal response to an increase in the acid load.
In the thick ascending limb of Henle’s loop, luminal NH4+ is reabsorbed by substitution for K, on the Na+>K,>2CΓ cotransporter. The NH4+ reabsorption in this segment reduces the amount of ammonia species delivered to the late distal tubule and increases ammonia (NH5) in the medullary interstitium.
High NH5∕NH4+ concentrations are enhanced and maintained in the medullary interstitium by a countercurrent multiplication system in the loops of Henle, similar to that described in Chapter 43. This creates a steep concentration gradient for NH5, which favors its movement into the medullary collecting duct. The established paradigm is that ammonia diffuses across plasma membranes and into the luminal fluid, where it acts as a buffer and combines with H' to form NH44. Because NH4 f is lipid insoluble, it cannot diffuse back across the apical plasma membrane and is trapped within the tubule fluid.
The formation of NH4' from intraluminal NH5 and H+ lowers the concentration of both NH5 and H+ in the tubule fluid. This contributes to the maintenance of a favorable gradient for the diffusion of NH3 into the tubule fluid and reduces the electrochemical gradient for H, that is created by active proton secretion in the collecting duct.
Until recently, NH5 diffusion and trapping in the collecting duct was believed to be the only significant mechanism of NH4* excretion. However, several studies have demonstrated transporter-mediated NH√ secretion in the collecting duct. Recently, members of the Rh glycoprotein family, which transport ammonium in other systems, have been found in the mammalian collecting duct. Although it remains to be proven that these transporters mediate ammonium secretion in the native collecting duct, in vitro studies suggest such a role for them.
Finally, the comparative aspects of ammonia excretion are intriguing. It is known that ammoniagenesis and ammonia excretion is an important mechanism controlling acid-base homeostasis in mice, rats, dogs, chickens, and humans. In these species, ammonia excretion accounts for up to 60% of net acid excretion in basal conditions and can increase to 90% of net acid excretion in models of metabolic acidosis. However, the findings in these animals cannot be applied generally to other species. Rabbits have low urinary ammonia excretion rates and do not increase ammonia excretion during metabolic acidosis. Some might not consider this surprising because rabbits normally excrete neutral or alkaline urine and might not be expected to have a full complement of mechanisms to excrete acid urine. However, domestic cats, which usually excrete acid urine, also do not increase renal ammonium excretion in response to a model of metabolic acidosis.
the filtered HCO5, it cannot maintain a large pH gradient across the apical plasma membrane. The net secretion of H+ in this segment is particularly dependent on the intraluminal buffers discussed earlier, which combine with secreted H4 and prevent the concentration of H, in the tubule fluid from rising significantly. As a result, although the majority of renal acid secretion (HCO3 reabsorption) occurs in the proximal tubule, the pH of the tubule fluid when it leaves this segment is similar to that of the glomerular filtrate.