The Lungs
The lungs are relatively small, unlobed, bright pink, and nonexpansile. Although somewhat firmer than mammalian lungs because they contain far more cartilage, the lungs of birds are soft and velvety to the touch.
They are confined to the craniodorsal part of the body cavity; their convex dorsal surface lies against and is deeply indented by the thoracic vertebrae and vertebral ribs. The lungs fail to cover the lateral surfaces of the heart as they do in mammals. The concave ventral (septal) surface lies against the horizontal septum (see further on) and faces the esophagus, heart, and liver (Fig. 37.27). The lungs are lightly attached to the body wall and to the horizontal septum that confines them from below. No pleural cavity corresponding to that of mammals is necessary because the capacity for expansion is negligible. The nonexpansile nature of the lungs, their abundant cartilage, and their confinement high within the body cavity surrounded by bone render them largely incompressible.The primary bronchus (Fig. 37.28) enters the ventral surface, passes diagonally through the lung (as the mesobronchus), narrowing as it goes, and at the caudal border becomes continuous with the abdominal air sac (Fig. 37.28/13; see later). In the chicken it gives off 40 to 50 secondary bronchi classified as medioventral, mediodorsal, lateroventral, and laterodorsal according to the general areas of the lung they supply (Figs. 37.27/a-d and 37.28/3-5). These groups of secondary bronchi have various connections with the air sacs; these communications are essential to the passage of air through the lungs.
The secondary bronchi give off 400 to 500 parabronchi, which contain gas exchange sites in their relatively thick walls. The parabronchi arise from the medioventral and mediodorsal bronchi and connect with each other end-to-end to form loops of various lengths (Fig.
37.28/6). These loops, which are tightly packed and parallel, constitute about three-quarters of the lung tissue, forming the functional division known as the paleopulmo. The parabronchi from the smaller lateroventral and laterodorsal bronchi form the less regular and more caudal functional division known as the neopulmo.The internal and external diameters of the parabronchi measure about 1 mm and 2 mm, respectively. The parabronchi anastomose with their neighbors, from which they are separated by fenestrated septa (Fig. 37.28//). Numerous extensions (atria) of the parabronchial lumen give rise to the air capillaries. These form a dense network of interconnected loops (Fig. 37.28/e) that spread into the interparabronchial septa. Anastomoses with air capillaries of adjacent parabronchi are found where the septa are deficient (Fig. 37.28/g). The air capillaries are closely intertwined with blood capillaries; the two networks constitute the bulk of the parabronchial wall. The arrangement of flow in the blood capillaries is crosscurrent, a feature contributing to the extreme efficiency of the avian lung. The air capillaries, about 5 μm in diameter, are lined by a single layer of epithelial cells resting on a basement membrane. The capillary endothelium is applied to the other side of the basement membrane. Gas exchange takes place across the barrier. The air capillaries are therefore comparable with the alveoli of the mammalian lung; the essential difference is that the air capillaries are not terminations of the respiratory tree but continuous channels that can receive oxygen-rich air from either direction.
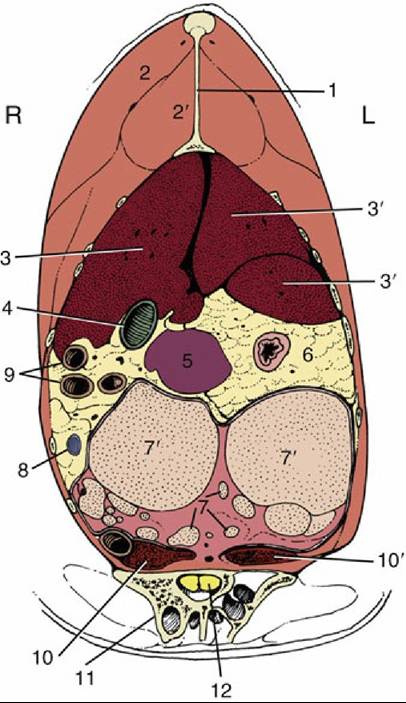
FIG. 37.24 Transverse section of the trunk at the cranial end of the ilium. L, Left; R, right; 1, keel of sternum, 2, pectoralis; 2', supracoracoideus; 3 and 3', right and left lobes of liver, respectively; 4, gallbladder; 5, spleen; 6, constriction between proventriculus and gizzard; 7, ovary; 7', follicle; 8, cranial mesenteric vein in mesenteric fat; 9, small intestine; 10 and 10', right and left kidneys, respectively; 11, ilium; 12, spinal cord.
Gas Exchange: Compared with mammals, the capture of oxygen in birds is much more efficient because of the following modifications: a thin blood-gas barrier, crosscurrent blood flow, oneway air flow, and pulmonary rigidity. However, the efficiency of gas exchange has its downside in that it makes birds much more susceptible to inhaled toxins and infections.
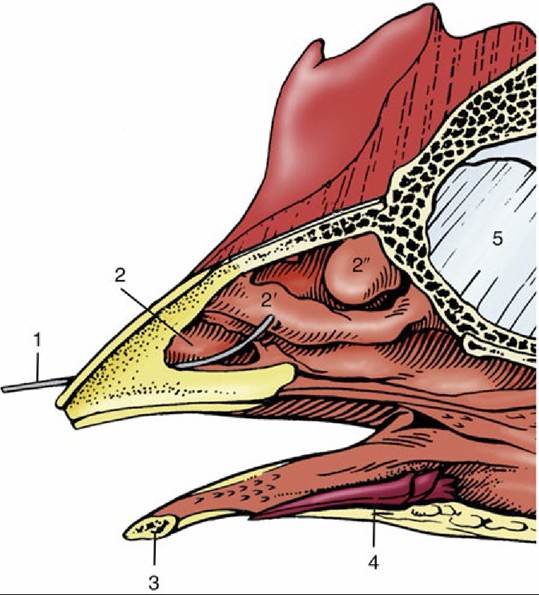
FIG. 37.25 Section through rostral part of the head of a chicken. 1, Wire in nostril; 2, 2', and 2", rostral, middle, and caudal nasal conchae, respectively; 3, mandible; 4, tongue; 5, interorbital septum.
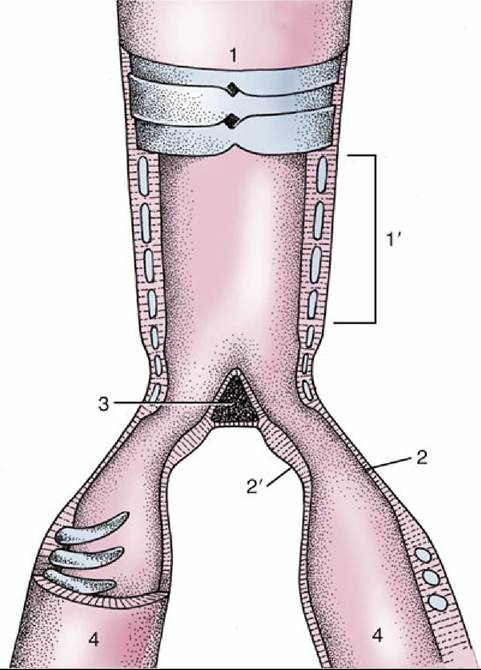
FIG. 37.26 Semi-schematic representation of the opened syrinx. 1, Trachea; 1', tympanum; 2 and 2', lateral and medial tympaniform membranes; 3, pessulus; 4, primary bronchi.
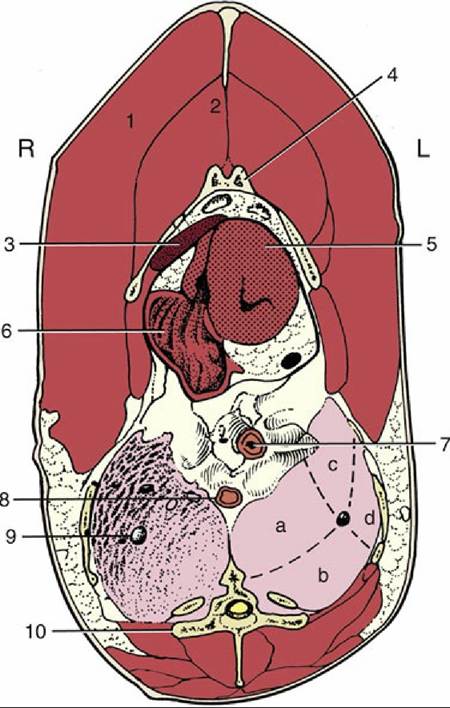
FIG. 37.27 Transverse section of the trunk at the level of the heart and lungs. L, Left; R, right; 1, pectoralis; 2, supracoracoideus; 3, liver; 4, sternum; 5, left ventricle; 6, right atrium; 7, esophagus; 8, descending aorta; 9, primary bronchus in right lung; 10, thoracic vertebra (notarium); a, b, c, and d, areas of left lung supplied by medioventral, mediodorsal, lateroventral, and laterodorsal secondary bronchi, respectively.
The air sacs are blind, thin-walled (two cells thick) enlargements of the bronchial system that extend beyond the lung in close relationship to the thoracic and abdominal viscera. Diverticula from some sacs enter various bones and even reach between muscles.
The chicken has eight air sacs: single cervical and clavicular, and paired cranial thoracic, caudal thoracic, and abdominal sacs. The cervical sac (Fig. 37.28/9) consists of a small central chamber ventral to the lungs from which long diverticula extend into and alongside the cervical and thoracic vertebrae.
The much larger clavicular sac lies in the thoracic inlet. Its thoracic part (Fig. 37.28/10') fills the space cranial to and around the heart and extends into the sternum; extrathoracic diverticula (Fig. 37.28/10) pass between the muscles and bones of the shoulder girdle to pneumatize the humerus. Compound fractures of the humerus may therefore introduce infection to the air sacs and lungs. The paired cranial thoracic sacs (Fig. 37.28/11) lie ventral to the lungs between the sternal ribs and the heart and liver. The paired caudal thoracic sacs (Fig. 37.28/12) lie more caudally between the body wall and the abdominal sacs. The paired abdominal sacs (Fig. 37.28/13) are the largest. They occupy the caudodorsal parts of the abdominal cavity, where they are in broad contact with the intestines, gizzard, genital organs, and kidneys. Their diverticula enter recesses of the synsacrum and the acetabulum.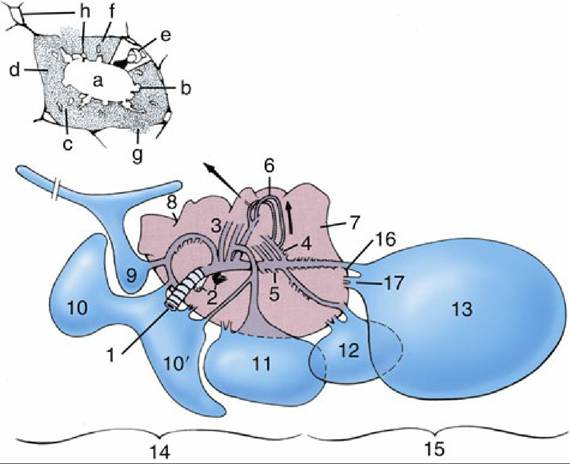
FIG. 37.28 Right lung (medioventral view) and related air sacs (schematic). The intrapulmonic structures have been simplified. 1, Primary bronchus; 2, pulmonary vessels at hilus; 3, medioventral bronchi; 4, mediodorsal bronchi; 5, lateroventral bronchi; 6, loops of parabronchi; 7, lung; 8, indentations caused by ribs; 9, cervical air sac; 10 and 10', extrathoracic and intrathoracic parts of clavicular air sac, respectively;
11, cranial thoracic air sac; 12, caudal thoracic air sac; 13, abdominal air sac; 14, cranial air sacs, functionally related to paleopulmonic parabronchi; 15, caudal air sacs, functionally related to neopulmonic parabronchi; 16, direct (sacculobronchial) connection of air sac to lung; 17, indirect (recurrent bronchial) connection of air sac to lung. Inset, Transverse section of a parabronchus. a, Lumen; b, atria; c, infundibula; d, network of air and blood capillaries; e, solidly drawn atrium and schematic air capillaries to show their continuity; f, interparabronchial septum; g, gas exchange tissue anastomosing through gap in interparabronchial septum; h, blood vessels.
The air sacs function primarily in respiration, although their poorly vascularized walls deny them any role in gas exchange. Nonetheless, healthy air sacs are requisite to normal lung function. Indeed, their general arrangement is such that, in stark contrast to the process in mammals, fresh air is moved through the lung on expiration as well as inspiration. This feature is an obvious contribution to the remarkable efficiency of the avian lung and the truly prodigious athletic capabilities it can support. The air sacs also lighten the body and, being largely dorsal, lower the center of gravity, presumably for improved stability in flight. Those in the body cavity sharply delineate certain organs in radiographs.
The cervical, clavicular, and cranial thoracic sacs form one (cranial) functional group connected to the ventral bronchi, and the caudal thoracic and abdominal sacs form a second (caudal) group connected to the primary bronchus. The cranial air sacs are thus related to the paleopulmo, the caudal to the neopulmo.
In summary, the air sacs function to create a unidirectional flow of air through the lungs, which is important for maximizing oxygen extraction. By evaporation, they also help to reduce the amount of heat produced during flight and may have subsidiary roles in sound production, courtship displays, and possibly cooling of the testes.
The account of respiration given here is greatly simplified. Inspiratory movements (in which the ribs are drawn forward and the sternum lowered) draw air through the lungs into the air sacs; the caudal sacs (Fig. 37.28/15) receive relatively fresh air, and the cranial sacs (Fig. 37.28/14) receive air that has already lost much oxygen by passing through the paleopulmonic parabronchi. On expiration the air sacs are compressed; most air from the caudal sacs now passes through the neopulmonic parabronchi, while most of that from the cranial sacs leaves through the trachea. The air sacs thus act like bellows, moving air through largely passive lungs.
The flow is circular, with air passing through the paleopulmonic parabronchial loops in the same direction. The feature is unique among vertebrates and in sharp contrast to the tidal flow in mammals.Respiration is effected by the intercostal and abdominal muscles. For inspiration there are six muscles, but the principal ones are the external intercostals and the costosternalis. On expiration the rib cage moves outward and the sternum moves downward. This negative pressure causes air to be sucked in via the nares to the air sacs. There are nine muscles of expiration, but the internal intercostal and the abdominal musculature are the main ones. On expiration the rib cage moves upward, which reduces the chest size and compresses air from the air sacs back through the lungs.
The avian flow-through system means that artificial ventilation can be achieved by passing fresh oxygen down the trachea or via an air sac cannula if the trachea is obstructed. Manual ventilation by compressing and lifting the sternum can also be undertaken.