The Mammary Glands
Mammary glands (mammae) are greatly modified, much enlarged sweat glands whose secretion nourishes the young. The modified milk (colostrum) produced immediately after parturition transfers immunoglobulins to the newborn.
Each mammary gland is a compound tubuloalveolar gland that consists of secretory units grouped into lobules defined by intervening connective tissue septa (see Chapter 29). The mammary glands develop as epithelial buds that grow into the underlying mesenchyme from linear ectodermal thickenings (mammary ridges). These ridges may extend from the axilla to the groin (as in carnivores and pigs) or may be of more limited extent, restricted to the axilla (as in elephants), the thorax (as in women), or the groin (as in ruminants and horses). Usually more buds appear than survive in the adult, and although most extra buds soon regress, some persist to give rise to supernumerary teats. These teats may be independent or may be attached to other, better developed glands (see Fig. 10.33A/7). They are unsightly, and because they may interfere with milking, they are often removed from the udders of cows and goats.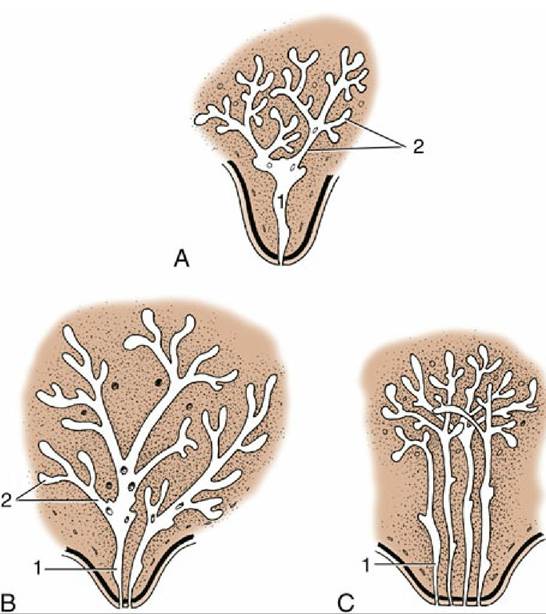
FIG. 10.31 Developing duct systems growing proximally from the tip of the fetal teat. (A) Cow, ewe, and goat. (B) Mare and sow. (C) Bitch and cat (only four primary sprouts are shown). 1, Primary sprout, which gives rise to the lactiferous sinus; 2, secondary and tertiary sprouts, which give rise to the lactiferous ducts.
Proliferation of the mesenchyme surrounding the bud raises a teat (papilla) on the surface of the body. One or more epidermal sprouts grow from the mammary bud into the connective tissue of the teat and begin to canalize at about the time of birth. Each sprout is destined to form a separate duct system with associated glandular tissue.
The mammary gland arising from only one sprout has a single duct system leading to a single orifice on the tip of the teat (Fig. 10.31A).The number of sprouts results in a corresponding number of separate duct systems, each with an associated glandular mass and separate orifice. The growth of the ducts and gland tissue is continued after puberty and especially during the first pregnancy, forming the swelling that pushes the teat away from the body wall. The process is controlled by the intricate interplay of several hormones from the hypophysis, ovaries, and other endocrine glands.
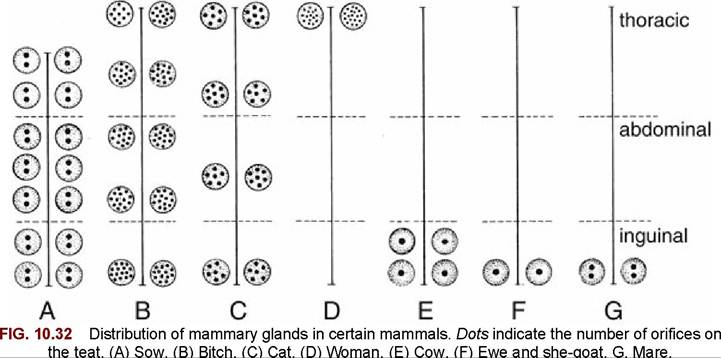
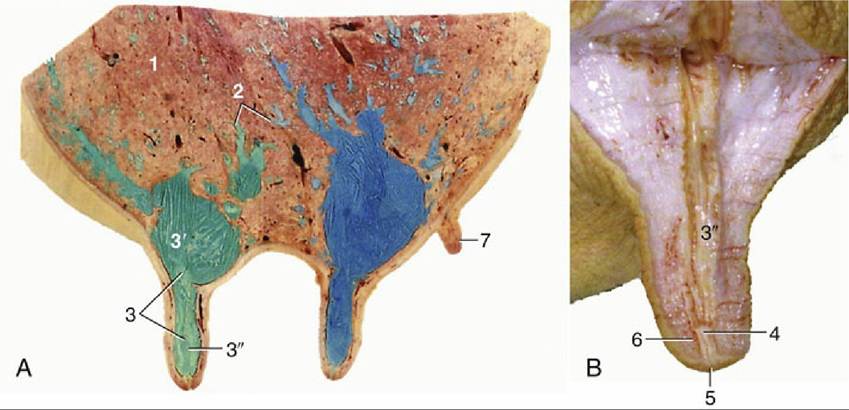
FIG. 10.33 (A) Sagittal section of udder, showing teat and gland sinuses and lactiferous ducts filled with latex (cranial quarter, green; caudal quarter, blue). (B) Section of teat. 1, Parenchyma of gland; 2, lactiferous ducts of various diameters; 3, lactiferous sinus; 3', gland sinus; 3", teat sinus; 4, papillary duct; 5, teat orifice; 6, teat sphincter; 7, supernumerary teat.
Each of the units formed along the trunk of a lactating sow (see Fig. 10.31B) is composed of glandular tissue supported and enclosed by a fibrous tissue framework in which the mammary vessels and nerves run. The whole formation is pervaded with fat and covered by skin. The mammary glands in species such as ruminants and horses may appear to merge in a single consolidated complex called the udder. This term is also applied to the even more distinctly separated mammary glands in the sow. The numbers of mammary glands (as well as their duct systems) in the domestic species is shown schematically in Fig. 10.32.
The more detailed organization is illustrated by reference to the cow. The glandular tissue is arranged in lobules, each 1 mm or perhaps a little more in diameter and consisting of about 200 alveoli. The milk drains to an intralobular duct that joins others to form a larger interlobular duct (Fig.
10.33/2). Interlobular ducts lead in their turn to a system of lactiferous (milk-carrying) ducts that ultimately convey the milk to the relatively large cavity known as the lactiferous sinus (Fig. 10.33/3). The lactiferous ducts of successive orders increase in diameter but diminish in number so that only 10 or so enter the sinus. Unlike most ducts, they have alternating narrow and dilated portions where contraction of the muscular wall of the narrow portions holds the milk in the expansions before it is "let down" when the cow suckles or is milked. The lactiferous sinus extends into the teat and is incompletely divided into gland and teat sinuses (Fig. 10.33/3' and 3 ") by a constriction. The teat sinus is continued by the papillary duct (Fig. 10.33/4), which opens at the tip of the teat, where the orifice is surrounded by a smooth muscle sphincter (Fig. 10.33/6).Corresponding parts can be identified in other species, including those in which each gland contains several small lactiferous sinuses, each served by a separate duct system and each opening independently.
It must be stressed that mammary glands are fully developed and fully functional only at the height of lactation. They are then large and show a predominance of yellow glandular tissue over the paler fibrous stroma. When the dam weans her young, involution sets in, the parenchyma regresses (see Chapter 29), and the connective tissues now form the bulk of the organ. However, the gland never quite reverts to its prelactation size, and it grows a little more with each pregnancy.
Mammary buds also form in male embryos and persist to give rise to the rudimentary teats found on the ventral surface of the trunk (carnivores and pig) or on the cranial surface of the scrotum (ruminants). They are less common in horses but occasionally appear beside the prepuce. In certain species such as rats, however, the male glands regress completely.
Comprehension Check
Demonstrate your understanding of the structure of the skin, its functions, and comparative anatomy of its modifications in the foot of the dog, cattle, and the horse.
* The coat of wild sheep and of extant primitive breeds exhibits an outer coat of very coarse, hollow-cored guard hairs, known as kemp, which conceals and protects, by facilitating the runoff of rain, a short undercoat of much finer wool fibers. The growth of both fiber types is seasonally restricted and is succeeded by a spring molt when the shed wool forms tangled mats that are eventually cast. The wool is harvested by being gathered from the pasture and plucked directly from the animals. Evolution of the fleece under domestication has been characterized by loss of pigmentation and by reduction in the amount of kemp, partly by depletion of the number of kemp hairs and partly by the transformation of a proportion of these into finer and more typical forms of hair. The wool now grows continuously and at a more rapid rate, though showing seasonal variation, and elimination of the spring molt introduces the necessity for shearing. The more rapid growth results in increased fiber length in the annual wool clip; other changes affect fiber waviness (crimp) and introduce greater diversity in the relative incidences of fibers of different diameters. The variations in these acquired features account for the characters, and therefore the values, of the fleeces of different breeds. The coarse, hairy fleece of some is most appropriate for the less valuable carpet trade, whereas the improved fleece of others is suited to the production of finer yarns and fabrics. The weight of wool produced annually also varies widely with breed, ranging from as little as 3 to as much as 20 pounds (1.4 to 9 kg).
* The pigment granules are produced within melanocytes, specialized cells of neural crest origin that are confined to the basal layer of the epidermis and hair follicles. The granules move to the tips of the dendritic processes of the melanocytes and are pinched off and subsequently phagocytized by neighboring cells (keratinocytes) in a process that continues until widespread. Melanin production is influenced by many factors including sufficiency of copper and, by the melanocyte-stimulating hormone. Melanin production may be affected by season (white fur in lagomorphs and mustelids in winter) and age.
* Formerly, and still occasionally, termed corium.
* Uniquely, the horns of the American Pronghorn are shed annually.
* There are important species differences. The distribution and other features of human (and other primate) sweat glands differ significantly.