The Most Important Body Pool of Calcium Involved in Homeostasis Is the Extracellular Fluid Component
Almost all calcium (99%) in the body is in bone in the form of hydroxyapatite crystals, which contain calcium, phosphate, and water. The next largest pool of calcium is intracellular calcium.
As stated previously, calcium is important for the response of cells in carrying out their physiological activities, including the secretion of hormones. In the inactive cell state, calcium concentrations are relatively low in the cytosol; calcium is bound to proteins or contained within the mitochondria or granules of the endoplasmic reticulum. Increased intracellular calcium concentrations are indicative of increased cell activity.The smallest pool of calcium, which resides in the extracellular Iluid (ECF), is the most important pool for physiological control of calcium concentrations in the blood. This component comprises interstitial calcium, blood calcium, and a small (0.5%) but important part of the bone calcium pool, which exists as amorphous crystals or in solution. The soluble bone calcium pool allows access to the large reserve of calcium that resides in bone.
The regulation of calcium levels involves control of the movement of calcium between the ECF and three body organs: bone, GI tract, and kidneys. The exchange of calcium ions between the ECF and intracellular fluid occurs in conjunction with the control of intracellular metabolism, with little effect on plasma concentrations of calcium.
The absorption of calcium from the Gl tract is by passive diffusion and active transport. The passive diffusion of calcium across the intestinal mucosa occurs in the presence of high concentrations and, as such, is not an important aspect of calcium absorption. Active transport involves the movement of calcium into the intestinal cell down a concentration gradient, which is facilitated by carrier proteins located on the luminal side of the mucosal cell.
Calcium is moved through the serosal side of the mucosal cell into the interstitial fluid through a calcium pump system. The active transport system adjusts according to the amount of calcium in the diet, becoming more active when calcium concentrations in the diet are lower and less active when calcium concentrations are higher. Calcium excretion into the GI tract is not affected by calcium uptake, and this can exacerbate conditions involving hypocalcemia. The Gl tract serves as the source of calcium for the body, even though both absorption and excretion of calcium occur through the tract. As discussed later, vitamin D plays an important role in the absorption of calcium from the GI tract.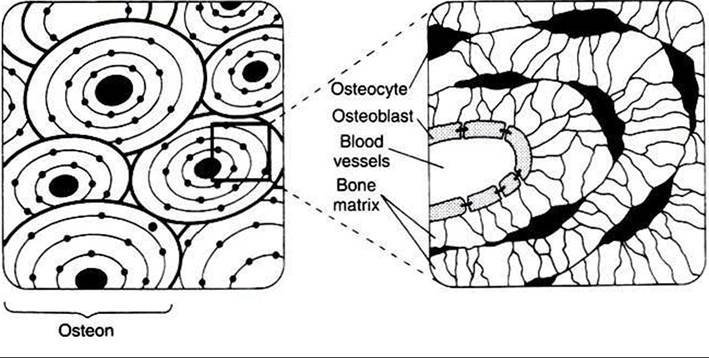
FIGURE 34-28 Structure of the osteon, the functional unit of bone, depicted in cross section at two magnifications. (From Hedge GA, Colby HD, Goodman RL: Clinical endocrine physiology, Philadelphia, 1987, Saunders.)
The kidneys serve as the route of excretion of calcium. Most of the calcium that passes into the kidneys is reabsorbed, with a net loss of only about 2%. This amount is matched by net absorption of calcium by the GI tract. Most of the calcium filtered by the kidneys is reabsorbed in the proximal tubules; the next largest amount is absorbed by the distal tubules, and a lesser amount, by the ascending loop of Henle. The distal tubules are under hormonal control and therefore are the sites of regulation of calcium in the kidneys.
The most important regulation of calcium metabolism between bone and ECF involves the soluble portion of bone. Amorphous crystals and soluble calcium, which form the source of ready exchange of ions with the blood, are located between the OSteoblastsf which line the blood vessel channels, and the osteocytes, which are deeper in the bone (Figure 34-28). These two cell types have cytoplasmic projections that interact intimately through the presence of tight cell junctions.
For labile bone calcium to reach the blood, calcium must cross the membrane barrier created by the osteoblasts and osteocytes. Movement of calcium from stable bone into the ECF also occurs but has little impact on the acute regulation of calcium concentrations. The process of remodeling bone, which occurs on a continuous basis, involves the breakdown Ofhydroxyapatite crystals by osteoclasts, a laying down of organic matrix by osteoblasts in the tunnels made by the osteoclasts, and finally the mineralization of the organic matrix by hydroxyapatite crystals. If an animal is subjected to prolonged changes involving calcium metabolism, the slowness of bone calcium exchange can have a significant impact on calcium metabolism.Parathyroid Hormone
The main organ involved in the control of calcium and phosphate metabolism is the parathyroid gland (Figure 34-29). Most domestic animals have four pairs of parathyroid glands that are generally located at the poles of the two lobes of the thyroid gland; the pig has only one pair of parathyroid glands, which lie anterior to the thyroid. The cranial pair of parathyroid glands in dogs and cats are at the Craniolateral poles of the thyroid, and those of ruminants and horses are anterior to the thyroid. The caudal pair of parathyroid glands in dogs, cats, and ruminants are located within the medial surface of the thyroid, whereas in the horse they lie near the bifurcation of the carotid trunk. I he parathyroid cells that are in the active process of hormone secretion arc called chief cells, whereas inactive, or degenerate, cells are called oxyphil cells.
The synthesis of parathyroid hormone (PTH) is similar to the synthesis of other protein hormones; a prepro-PTH of 115 amino acids is synthesized in the rough endoplasmic reticulum and then cleaved by 25 amino acids to form pro-PTH. A 6-amino acid “pro" portion is removed by the Golgi apparatus; the resulting PTH has 84 amino acids. PTH is secreted by the process of exocytosis.
PTH is rapidly metabolized by the liver and kidneys and has a relatively short half-life (5-10 minutes) in blood.The effect of PTH is to increase calcium and decrease phosphate concentrations in ECFs. PTH has direct effects on bone
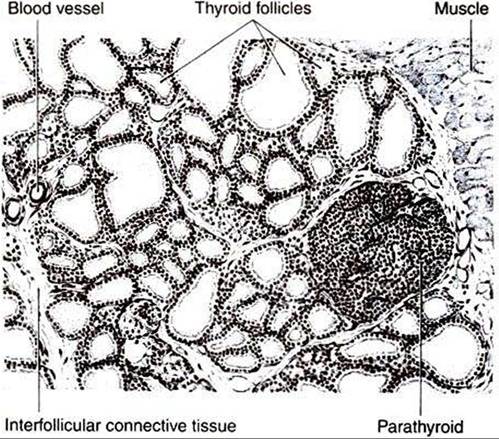
FIGURE 34-29 Depiction of a section of the thyroid and parathyroid glands of the rat as seen under low power of the microscope. Notice that the parathyroid gland lies near the surface of the thyroid gland and is surrounded on three sides by the thyroid follicles. (FromTurner CD, Bagnara JT: General endocrinology, ed 6, Philadelphia, 1976, Saunders.)
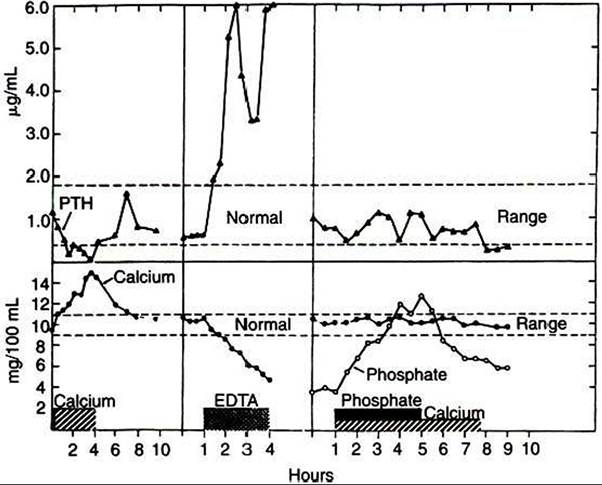
FIGURE 34-30 ■ Changes in plasma immunoreactive parathyroid hormone (PTH) levels in response to hypercalcemia induced by calcium infusion; in response to hypocalcemia produced by ethylenediaminetetraacetic acid (EDTA) infusion; and in response to hyperphosphatemia with normocalcemia in a cow. (From Capen CC: The calcium regulating hormones: parathyroid hormone, calcitonin, and Cholecalciferol. In McDonald LE, Pineda MH, editors: Veterinary endocrinology and reproduction, Philadelphia, 1989, Lea & Febiger.)
and kidney metabolism of calcium and indirect effects on GI metabolism of calcium. The initial effect of PTH on bone is to promote the transfer of calcium across the osteoblast-osteocyte membrane. This level of action occurs without the movement of phosphate and therefore has no effect on phosphate concentrations in blood. PTH has additional effects on stable bone, which results in the resorption of the bone. This effect involves increased osteoclast activity and an inhibition of osteoblast activity. The effect of PTFl on stable bone results in the release of both calcium and phosphate.
PTH acts on the distal convoluted tubules of the kidneys to increase absorption of calcium and decrease renal phosphate reabsorption through an effect on the proximal tubules.
PTH is involved also in the activation of vitamin D at the kidney level. PTH mediates the absorption of calcium from the gut indirectly through its effect on vitamin D.PTH secretion is controlled by free (ionized) calcium concentrations in blood; decreases in calcium levels stimulate PTH secretion, and increases in calcium turn off secretion (Figure 34-30). Both actions are mediated by an effect on cAMP metabolism. Epinephrine stimulates PTH secretion through stimulation of β-adrenergic receptors. Magnesium affects PTH secretion in the same manner as calcium, but its physiological impact is much less. Sleep affects the secretion of PTH; values are highest immediately after waking.
Calcitonin
Calcitonin, a hormone produced by cells in the thyroid gland, also affects calcium metabolism. Cells of the type involved in the synthesis of calcitonin—parafollicular, or C, cells—are scattered throughout the thyroid gland and are distinctly different from the cells that synthesize thyroid hormones. During the early studies of calcitonin in animal classes such as fish, amphibians, reptiles, and birds, which have separate thyroid and Ultimobranchial glands, it was found that all the calcitonin activity was in the Ultimobranchial glands. Therefore the calcitonin cells represent Ultimobranchial gland tissue that has been incorporated into the thyroid during embryonic development.
Calcitonin, synthesized as a preprohormone, has 32 amino acids; a ring structure at the amino terminus contains a disulfide link that bridges between amino acids I and 7. The processing of the molecule is interesting because calcitonin is located in the middle of procalcitonin, so an additional enzyme cleavage is required for the formation of the active molecule. The secretion of calcitonin is by exocytosis from granules.
Calcitonin acts as a counterbalance to PTH because it causes hypocalcemia and hypophosphatemia. The effect of calcitonin on mineral metabolism is mainly on bone (Figure 34-31).
Calcitonin decreases the movement of calcium from the labile bone calcium pool (behind the osteoblast-osteocyte barrier) to the ECF and decreases bone resorption through an inhibitory effect on osteoclasts. Whereas the inhibition of bone resorption explains one aspect of the hypophosphatemic effects of calcitonin, calcitonin also increases movement of phosphate from the ECF into bone. Calcitonin decreases Gl activity directly by inhibiting gastric acid secretion and indirectly by inhibiting gastrin secretion. The physiological importance of this is not known. Calcitonin also increases renal excretion of calcium and phosphate.The control of calcitonin secretion is by calcium; increased calcium concentrations cause increased secretion of calcitonin. The physiological control of calcium metabolism by calcitonin operates in situations of hypercalcemia with increased secretion of calcitonin and concomitant inhibition of PTH secretion. During hypocalcemic conditions (Figure 34-31), calcitonin synthesis is inhibited, and PTH becomes responsible for reestablishing normal calcium concentrations in the ECFs. GI hormones, including gastrin, cholecystokinin,
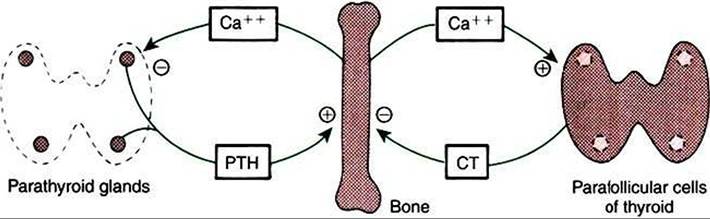
FIGURE 34-31 Negative-feedback loops controlling parathyroid hormone (PTH) and calcitonin (CT) secretion. Plus signs indicate stimulation; minus signs indicate inhibition. (From Hedge GA, Colby HD, Goodman RL: Clinical endocrine physiology, Philadelphia, 1987, Saunders.)
secretin, and glucagon, stimulate the secretion of calcitonin, with gastrin the most potent. These hormones limit postprandial hypercalcemia.
Vitamin D
Vitamin D is important for the absorption of calcium from the gut. It is a Steroidlike molecule, and because it is produced in one tissue and transported by the blood to a distant site of action, it should probably be called a hormone instead of a vitamin. All the vitamin D produced by the body is produced in the skin. Epithelial cells of the skin synthesize the immediate precursor of vitamin D, 7-dehydrocholesterol, from acetate. Exposure of the skin to ultraviolet light results in cleavage of the C-9 and C-IO bonds of 7-dehydrocholesterol, which results in the formation of vitamin D (Figure 34-32). The vitamin D molecule, as such, is inactive and must be transformed by both the liver and the kidney before the molecule is biologically activated. The liver first hydroxylates the molecule at the C-25 position, and the kidney subsequently hydroxylates the molecule at C-I to produce the active compound, 1,25- (OH)2-Vitamin D (1,25-vitamin D).
Control of the C-I hydroxylase in the kidney by PTH is the most important control linkage for the synthesis of 1,25- vitamin D. Decreases in calcium concentrations stimulate PTH secretion, which in turn favors the synthesis of active vitamin D and increased intestinal absorption of calcium. Phosphate also regulates vitamin D metabolism. Increased serum phosphate concentrations stimulate an enzyme that promotes hydroxylation of C-24 (instead of C-I) by the kidney, which leads to the formation of 24,25-(OH)2-vitamin D, an inactive molecule. The active molecule, 1,25-vitamin D, also regulates itself by decreasing C-I hydroxylase and increasing C-24 hydroxylase activity; decreased amounts of active vitamin D is the result.
Because of its lipid nature, 1,25-vitamin D is transported by binding to proteins in the plasma. Most of vitamin D is carried in association with a specific α globulin called trαns- calciferiny a molecule synthesized by the liver.
The most important effects of vitamin D involve increased absorption of calcium by the GI tract. Vitamin D stimulates the synthesis of protein within the mucosal cells, which aids the rate-limiting step in calcium absorption: movement of cal-
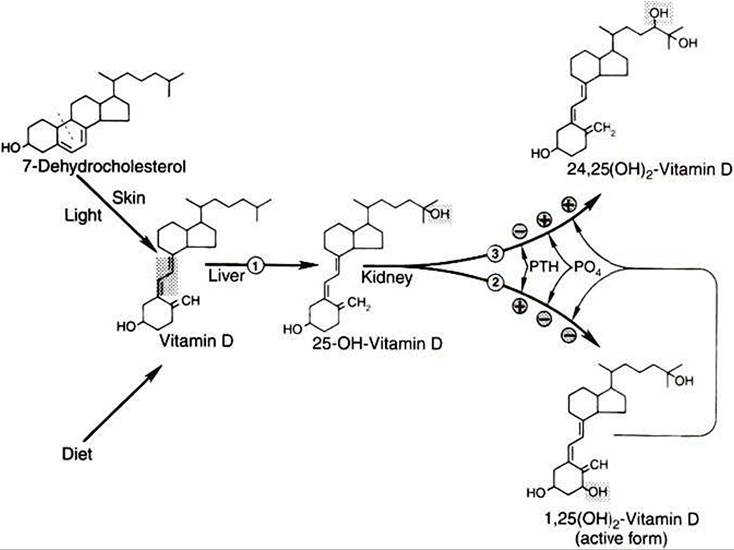
FIGURE 34-32 Synthesis and metabolism of vitamin D.The position of hydroxylation of 25-OH-vitamin D in the kidney is controlled by parathyroid hormone (PTH)t phosphate (PO4)1 and 1,25-(OH)2-vitamin D. Shading indicates structural change at each step; dashed line indicates the position of cleavage of 7-dehydrocholesterol to produce vitamin D. Enzymes:
(1) 25-hydro?ylase,
(2) 1α-hydroxylase,
(3) 24-hydroxylase. Plus signs indicate stimulation; minus signs indicate inhibition. (From Hedge GA, Colby HD, Goodman RL: Clinical endocrine physiology, Philadelphia, 1987, Saunders.)
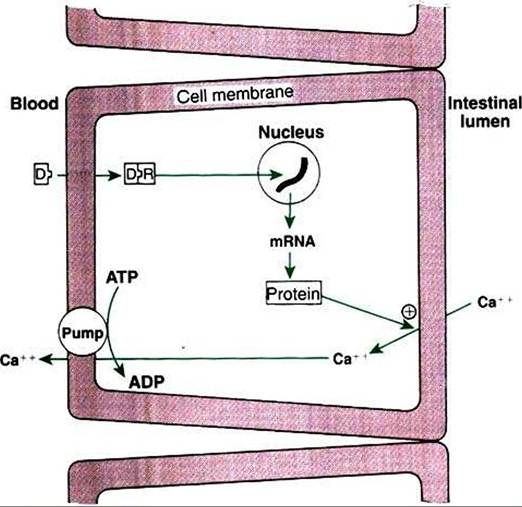
FIGURE 34-33 Mechanism of action of 1,25-(OH)2-vitamin D (D) to increase calcium absorption in the intestine. Plus sign indicates stimulation. R, receptor. (From Hedge GA, Colby HD, Goodman RL: Clinical endocrine physiology, Philadelphia, 1987, Saunders.)
cium into the mucosal cell (Figure 34-33). Because the intestinal effect of vitamin D depends on the activation of protein synthesis by mucosal cells, the effect on calcium absorption usually requires several hours. Although the stimulation of protein synthesis relates mostly to active transport of calcium, vitamin D also stimulates passive transfer of calcium. Vitamin D also has effects on bone, promoting the movement of calcium ions from the labile pool into ECFs and the resorption of bone, as well as enhancing the effects of PTH on bone metabolism of calcium.
Thecontrol of 1,25-vitamin I) synthesis is by PTH and phosphate. A decrease in calcium concentrations results in increased PTH secretion and increased formation of 1,25-vitamin D through enhancement of C-I hydroxylation (Figure 34-34). This action leads to the correction of hypocalcemia by increasing absorption of calcium by the gut. A decline in phosphate concentrations results in decreased inhibition of the C-I hydroxylation, which indirectly results in increased 1,25- vitamin D production and increased absorption of phosphate. Some evidence suggests that hormones associated with pregnancy, such as growth hormone and prolactin, increase 1,25- vitamin D production by stimulating C-I hydroxylation.
In the overall control of calcium metabolism, PTH is primarily responsible for the maintenance of calcium homeostasis. The primary target tissue for Pl H in calcium homeostasis is the labile pool in bone; changes in renal absorption of calcium are also important. In the case of long-term calcium deficit in the diet, both PTH and 1,25-vitamin D are important for correction of the deficit. Decreased dietary calcium leads to decreased concentrations of calcium in the ECFs and
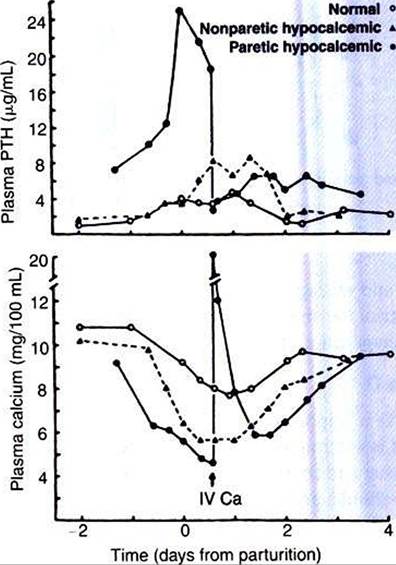
FIGURE 34-34 Development of varying degrees of hypocalcemia in cows near parturition, with corresponding increases in plasma parathyroid hormone (PTH) Ievels-The cow developing severe hypocalcemia (14 mg∕dL) can progress rapidly to acute renal failure when the calciumphosphorus product (Ca ? PO4) exceeds 60 to 80 mg/dL because of mineralization of renal tissue.
The diagnostic approach to hypercalcemia consists of ruling out the most common cause: hypercalcemia of malignancy. A thorough history and physical examination, including lymph node and rectal examination (for anal sac adenocarcinoma), complete blood cell count, urinalysis,serum chemistry profile, and chest and abdominal radiographs, are necessary to search for underlying neoplastic processes. Iflymphoma is not detected on the minimum database, a bone marrow examination and survey skeletal radiographs may be necessary. Once a diagnosis of neoplasia has been excluded, the next primary differential for hypercalcemia is chronic renal failure. This is the most difficult differential to exclude because other causes of hypercalcemia may result in renal damage because of soft tissue mineralization of the kidneys. Therefore an animal with hypercalcemia, azotemia, and hyperphosphatemia could have primary hyperparathyroidism, primary renal failure with secondary renal hyperparathyroidism, or vitamin D intoxication. Furthermore, patients with hypercalcemia secondary to renal disease may also exhibit elevations in intact PTH. Diagnosis of primary hyperparathyroidism is based on the findings of hypercalcemia (preferably ionized), hypophosphatemia (unless azotemic), high-normal to elevated serum PTH concentrations, and a mass in the cervical region. Intact PTH, demonstrated by a “sandwich** assay validated for use in the dog and cat, should be measured. A normal PTH concentration in the presence of elevated total and/or ionized calcium is considered inappropriate for the calcium level and would be considered diagnostic for primary hyperparathyroidism. For suspected cases of hypercalcemia of malignancy in which the diagnostic approach has failed to identify a neoplastic process, PTH- related protein (PTH-rp) concentrations may be measured.
The classic biochemical findings in animals with hypoparathyroidism are hypocalcemia (both total and ionized) and hyperphosphatemia. Other causes of hypocalcemia include iatrogenic (post-thyroidectomy) hypoparathyroidism, chronic and acute renal failure, acute pancreatitis, Iiypoalbuininemia, puerperal tetany (eclampsia), ethylene glycol intoxication, intestinal malabsorption, and nutritional secondary hyperparathyroidism. Early signs of hypocalcemia are nonspecific and include anorexia, facial rubbing, nervousness, and a stiff, stilted gait. Later signs progress to paresthesias, hyperventilation, and finally generalized tetany and seizures.
Primary hypoparathyroidism is diagnosed by means of an intact PTH assay. Serum or plasma PTH concentrations should be measured on a freshly drawn morning sample in a fasting animal. Handling of the sample is crucial to appropriate diagnosis because PTH may degrade if subjected to warm temperatures. Intact PTH refers to the entire 85-amino acid sequence of PTH; this is measured in a double-antibody “sandwich” assay in most endocrine laboratories that perform PTH measurement. For the diagnosis of primary hypoparathyroidism, the sample should be analyzed for both ionized calcium and intact PTH. Low ionized calcium and undetectable intact PTH concentrations are diagnostic for hypoparathyroidism.