THE OVIDUCT OR THE FALLOPIAN TUBE
The ovary and uterus are joined by an oviduct. Its four functional divisions are the isthmus, ampulla, infundibulum, and fimbriae (Figure 21.6). The oviduct’s ovarian end enlarges to form fimbriae, which have finger-like projections or fringed edges.
The ovarian bursa is made up of fibriae and a thin meso-salpinx peritoneal fold. In farm animals, the ovarian bursa is typically open, but in pigs, it covers the entire ovary. Only the ovulation fossa is covered by the bursa in mares. During ovulation, free-moving fimbriae sweep over the ovarian surface. The second portion of the oviduct, known as the infundibulum, is shaped like a funnel and presents the ostium abdominale, the oviduct’s abdominal opening. The animal’s age and size affect the infundibulum’s dimensions. Ampulla joins the infundibulum to the isthmus and makes up more than half of the oviduct’s length. At the utero-tubal junction (UTJ), the isthmus and uterus are directly connected.Both inner circular and outer longitudinal smooth muscles are present in the oviduct walls. From the ovarian to
TABLE 21.1
The Functions of Various Parts of Duct System
Oviduct Ideal conditions for preimplantation growth and fertilization
Uterus Early embryogenesis, attachment of the conceptus, and fetus expulsion through fetal membranes.
Cervix Barrier; during estrus, secretes mucus; during pregnancy, the cervical seal is produced.
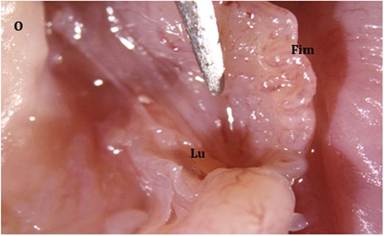
FIGURE 21.6 Gross morphology of the oviduct showing fimbriated end and lumen in cats: Ovary (O), Fimbriae (Fim), & Lumen of oviduct (Lu).
the uterine end, the muscle thickness increases. The oviduct gets shorter when muscles contract longitudinally, and annular constriction occurs when muscles contract circularly. Three different types of contractions are seen in oviducal musculature, viz., the localized peristalsis and segmental contractions.
Due to the possibility of simultaneous contractions or specific patterns, the oviduct’s motility is complex. Different oviduct segments also have different patterns and amplitudes of contractions. During ovulation, oviductal muscle contractions peak in intensity and complexity before gradually declining. The primary, secondary, and tertiary folds comprise the oviducal mucosa.
The mucosa in the ampulla is arranged into high- branched folds that become low ridges at the UTJ and get shorter as they approach the isthmus. At the ovarian end of the oviduct, the lumen is entirely filled with these folds.
One layer of columnar cells, both ciliated and non-cil- iated in type, makes up the mucosa (Figures 21.7 & 21.8). Hormonal activity affects the ciliary beat. During ovulation, the movement of fimbriae and this ciliary beat are synchronized. These cilia beat at their fastest during ovulation and then gradually slow down afterward. Normally, these cilia beat in the direction of the uterus. Along the oviduct, the concentration of ciliated and non-ciliated cells varies, most likely in relation to its function. For instance, the ciliated cells are most abundant at the fimbriated end, where their primary role is to pick up the ovum. On the other hand, the non-ciliated secretory cells (Figure 21.7) are most abundant in the ampulla and other areas where fluids are required to support gamete interaction and embryonic nourishment.
The nature of non-ciliated cells is secretory. They have secretory granules with many microvilli covering their apical surface. The oviductal lumen receives the release of secretory granules that accumulate in the epithelial cells during the follicular phase of the estrous cycle, thereby reducing their height. Oviductal fluid flows in the direction of the ovary. For two to three days following estrus, the majority of it is lost into the peritoneal cavity. However, it then descends the tube into the uterus, where it carries the
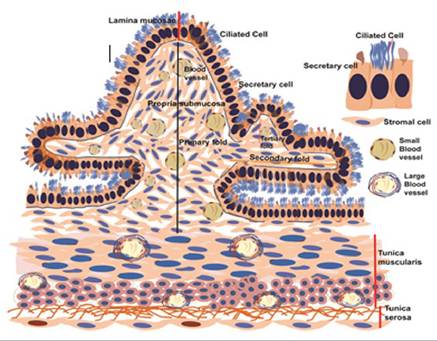
FIGURE 21.7 Schematic diagram of various histologic layers of oviduct.
embryos for implantation. The functional activity of ciliated and secretory cells in the oviduct is reliant on ovarian estrogens. The oviduct is responsible for transporting spermatozoa and eggs in opposing directions at roughly the same time. These disparate activities never fail because of how well the tubes’ structure and function complement one another. The ovum is transferred from the ovarian surface into the infundibulum by the fimbriae. The delicate balance between the ciliary beating towards the ostium and the cumulus-oocyte complex’s adhesion to the infundibulum is what causes this ovum pickup. The extracellular matrix that is present between the cumulus cells that are attached to the ciliary tips of the fimbriae in the infundibulum facilitates the adhesion of the COC. It is possible for further fimbriae to absorb eggs shed from the contralateral ovary. Oviductal secretions serve several purposes: (i) they capacitate sperm; (ii) they create an environment that is favorable for fertilization; (iii) they nourish the developing embryo; (iv) they protect oocytes, sperm, and embryos from the mother’s immune system as foreign antigens; and (v) they cause zygote cleavage by secreting a particular protein that causes cleavage. The ciliary movement, in conjunction with fluid movement and muscular contractions, aids in the following processes: (i) oviductal contents mixing, which increases the likelihood of male and female gametes encountering each other; (ii) denudation of follicular cells adhering to the ovum during ovulation; (iii) zygote transport into the uterus; and (iv) prevention of tubal implantation, or ectopic pregnancy. Oviductal peristalsis does not speed up but rather delays the entry of early embryos into the uterus.
21.10