THE UTERUS
All domestic animals have a uterus, which is made up of a body and two horns. The uterus is characterized as bipartite in cows, ewes, and mares, with the horns fully divided by a septum with a separate uterine body.
This is substantial, particularly in the mare. The sow’s uterus is bicornuate, with a short body connected to extremely long (4-5 feet) folded horns. Anatomically speaking, mammals that bear children have long uterine horns. Other mammal uteri types include simplex (found in primates), duplex (found in rabbits and guinea pigs), and diadelphous (found in platypuses). In animals with diadelphous uteri, the male has a penis that is forked and enters both reproductive openings at the same time. It’s interesting to note that farm animals occasionally have primitive uterine structures like duplex or diadelphous, but these do not affect fertility.The uterus is made up of three distinct layers: the inner endometrium, which is made up of epithelium, glands, and connective tissue; the outer serous membrane, which is an extension of the peritoneum; and the middle myometrium, which is made up of outer longitudinal, inner circular smooth muscles and a vascular layer in between (Figures
21.9 & 21.10).
All domestic animals have simple columnar non-ciliated luminal epithelium in their uteri. During estrus, these cells are tall and actively secretory; however, over the next two days, they become low cuboidal. Estrogens therefore stimulate the endometrial epithelium. The uterine lumen is where the coiled and branching tubular structures known as endometrial glands open. At estrus, they are straight, but as their secretory activity rises during the luteal phase of the cycle, they become more coiled and complex. The endometrial
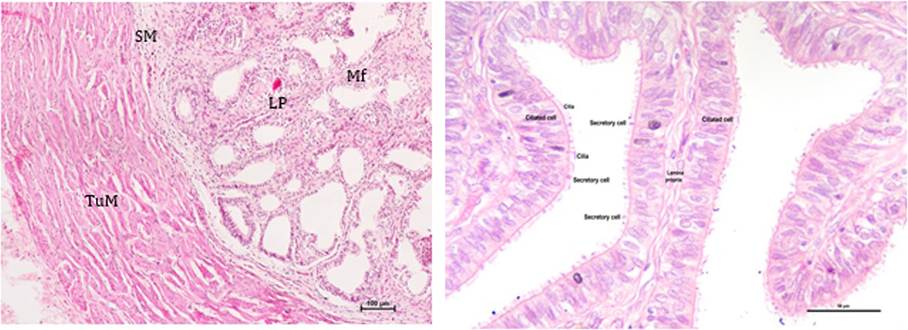
FIGURE 21.8 Photomicrograph showing various histologic layers of the oviduct (a) and the different cell types in the epithelium (b): Tunica muscularis (TuM), Submucosa (SM), Lamina propria (LP), and Mucosal folds (MF).
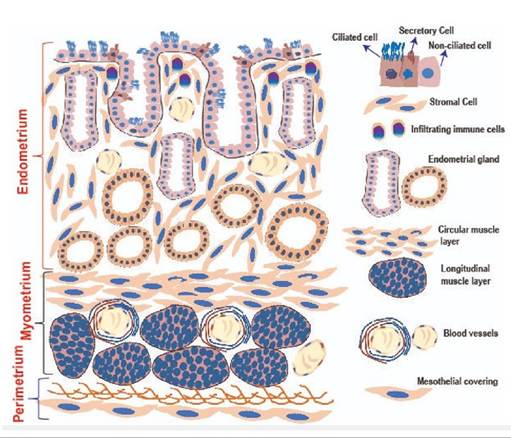
FIGURE 21.9 Schematic diagram of various histologic layers of the uterus.
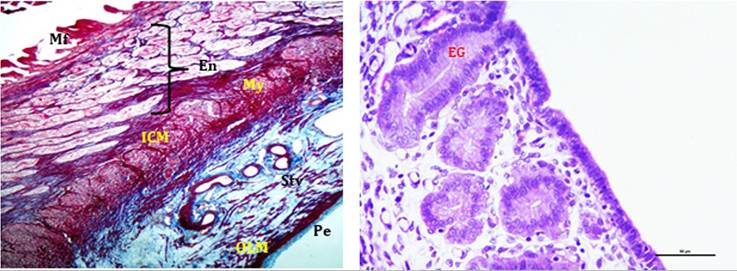
FIGURE 21.10 Photo micrograph showing various histologic layers of uterus (a) and uterine glands (b): Mucosal folds (Mf), Endometrium (En), Myometrium (My), Inner circular muscle layer (ICM), Outer longitudinal muscle layer (OLM), Stratum vasculosum (Stv), Perimetrium (Pe) & Endometrial glands (EG).
secretions’ volume and composition change according to the stage of the reproductive cycle. Various critical proteins, including uteroferrin, glyceryl phosphoryl choline diesterase (GPC), blastokinin, and implantation promoting factor, are detected in endometrial secretions during distinct stages of the estrous cycle. The ruminant endometrium has specific places where the fetal membranes can attach. “Cotyledons” are the elevated structures that make up these sites. At the cotyledon, endometrial glands are absent.
The regular motions of the oviduct are timed with the contractions of the myometrium. Generally, during estrus, the contractions travel towards the ovary; later, they move towards the cervix. During various stages of the estrous cycle, uterine contractility is altered by ovarian steroid hormones. Progesterone has the opposite effect of estradiol, which causes uterine contractions to occur more frequently.
Sperm must be transported from the site of ejaculation to the site of fertilization by means of uterine contractions that take place during mating. Therefore, it is thought that rubbing the uterus gently at AI will increase fertility. Following estrus, the uterine contractions move the zygote into the uterus for implantation as they are directed towards the cervix. Elevated progesterone levels prevent uterine contractions during the course of pregnancy. This progesterone-induced inhibition disappears as parturition gets closer, and the strong uterine contractions that ensue are essential for the fetus’s expulsion.
During estrus, endometrial secretions cause sperm to become capacitated, which is a necessary stage of maturation for fertilization. They also speed up the metabolism of sperm. In this regard, GPC plays a crucial role in releasing glycerol from phosphoglycerol so that sperm can use it. Preimplantation and early post-implantation embryos receive nourishment from endometrial secretions during the post-estrous/luteal phase. In farm animals, endometrial secretions appear to play a significant role in the nutrition of early post-implantation embryos. For instance, cow embryos do not firmly adhere to the endometrium; instead, they live freely and multiply widely for up to 30 days following the estrus. The sensitivity of the myometrium and the proliferative response of the endometrial epithelium and glands to ovarian steroids are modulated by endometrial protein secretions, particularly those of the estrogen and progesterone receptors. A particular protein called blastokinin is found in the uterine secretions of rabbits, and it helps morula stage embryos form blastocysts. Uterine secretions in mice contain an implantation-promoting factor. It is also thought that uteroferrin, which is present in ewes’ endometrial secretions, promotes embryonic development and implantation. Only the uterus could serve as a home for pregnancy. Ectopic pregnancy, or placement of the fetus elsewhere, is dangerous for the mother as well as the fetus. The uterus is uniquely adapted to experience profound alterations in dimensions, composition, and blood flow that are necessary to support the preimplantation embryo from the beginning of gestation to the end of parturition. The uterus is crucial to the upkeep or decline of CL. The uterus notifies the CL of the existence or lack of a conceptus by sending the proper signals. A functional CL lives longer after a hysterectomy, but a partial hysterectomy causes it to regress. In a similar vein, depending on when they are implanted, intrauterine devices may also lengthen or shorten CL’s life. The utero-ovarian veno-arterial pathway is a local pathway that carries these signals.21.11