The Rate of Gastric Emptying Must Match the Small Intestine's Rate of Digestion and Absorption
The rate al which food leaves the stomach must match the rate at which it can be digested and absorbed by the small intestine. Because some types of foods can be digested and absorbed more rapidly than others, the rate at which the stomach empties must be regulated by the contents of the small intestine.
Thus there are reflexes that regulate gastric emptying and allow the stomach to serve as a storage site. The afferent receptors of these reflexes are in the duodenum and are activated by low pH, high osmolality, and the presence of fat. Separate sensory receptors apparently exist for each of these stimuli, but these receptors have not been identified anatomically.Many reflexes occur within the Gl system. Their names usually reflect the site of origin of the afferent stimulus and the site of the efferent response. Thus, reflex control of gastric
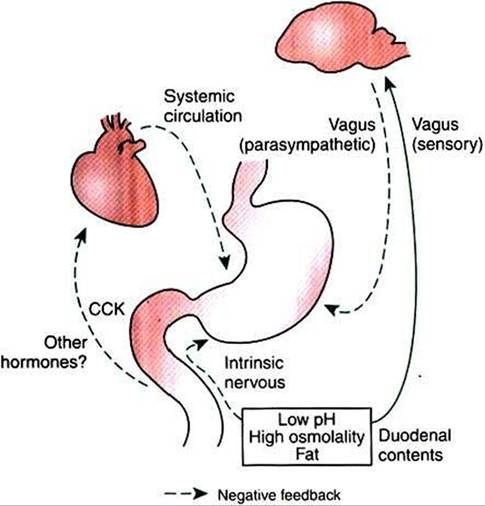
FIGURE 28-8 Inhibitory arcs of the enterogastric reflex. Low pH, high osmolality, and the presence of fat in the duodenum stimulate vagal, enteric neuronal, and hormonal reflexes that inhibit stomach emptying. After the duodenal pH and osmolality have moderated and some of the fat has been absorbed, the inhibitory influences on the stomach are removed. CCKf Cholecystokinin.
emptying by the duodenum is referred to as the enterogastric reflex (“entero” referring to the intestine).
The arc of the enterogastric reflex probably involves both the CNS and the ENS, as well as the endocrine/paracrine system (Figure 28-8). The extrinsic reflex pathway appears to involve afferent fibers of the vagus nerve, which receive stimuli in the duodenum. These stimuli are integrated in the brainstem, and the response is mediated by vagal efferent fibers to the stomach. The enteric reflex arc involves receptors in the duodenum and nerve fiber connections in the ENS that directly affect gastric emptying.
A contribution of the GI endocrine system to the enterogastric reflex has long been suspected, but the exact hormones responsible for the reflex are not known. CCK and secretin may be important. Both hormones are secreted by cells in the duodenum; CCK is secreted in response to fat and secretin in response to low pH; both appear to have suppression of gastric emptying as secondary effects. Gastric inhibitory peptide is a hormone produced in the duodenum in response to the presence of carbohydrate. In the dog, gastric inhibitory peptide may function as an inhibitor of gastric emptying, although stimulation of insulin secretion is probably its major action.
Enterogastric reflexes control gastric emptying by regulating stomach motility. The manner in which motility affects gastric emptying of solids is different from that for liquids. The rate at which solids are expelled from the stomach is regulated by the rate at which they are broken down into particles small enough to pass through the pylorus. This in turn is controlled by the motility of the antrum, or distal stomach; the greater the motility of the antrum, the faster material is broken down. Thus, antrum motility regulates the rate of release of solid material from the stomach. Liquid material leaves the stomach more quickly than solid matter, and the release of liquid may be less dependent on antral motility than on the motility of the proximal stomach.
Minimal mixing activity occurs in the proximal stomach. Therefore, liquids and solids tend to separate, the liquids moving to the outside and the solids to the center of the mass of food in the proximal stomach. Increased tension in the wall of the stomach body forces liquid into the antrum. Liquid may leave the antrum quickly, dependent on the activity of the pylorus. On the other hand, increased tension in the stomach body has little effect on the transport of solid material, because such material cannot leave the gastric body until sufficient space has been made available in the antrum.
Thus, motility of the stomach body appears primarily responsible for the liquid-emptying rate, whereas motility of the antrum is most responsible for the solid-emptying rate. The effect of the pylorus itself on gastric emptying is not as great as might be expected; removal of the pylorus results in a slight increase in the liquid-emptying rate and little increase in the rate of emptying of solid material. It appears that the distal portion of the antrum can account for much of the sieving action usually attributed to the pylorus. The rate of emptying of an isotonic liquid from the stomach is exponential and dependent on the initial volume of the liquid meal. Under usual circumstances, a liquid meal in the canine stomach has a half-life of about 18 minutes and is essentially gone by 1 hour after ingestion. Solid material is emptied more slowly, and its rate depends on its fat content. Low-fat meat meals are usually gone from the stomach in 3 to 4 hours after ingestion.