The Skull
The complete skeleton of the head comprises the skull,* the mandible or lower jawbone, the hyoid apparatus, the ossicles of the middle ear, and the cartilages of the external ear, nose, and larynx.
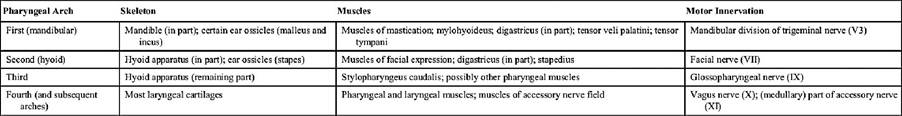
The skull (in the narrower sense) is a mosaic of many bones, mostly paired but some median and unpaired, that fit closely together to form a single rigid construction. The separate elements, which are named individually, develop from independent centers of ossification and have, for the most part, well-established homologies. In the young animal they are separated from each other by narrow strips of fibrous tissue — cartilage in a few situations — and this pattern of joints or sutures provides for growth. Once growth has ceased, ossification extends into the connective tissue, finally welding the bones together. This process is drawn out, and it may never be completed; the outlines of most bones are therefore discernible, even in skulls of old animals. Acquaintance with the names, positions, and approximate extents of the individual bones (Fig. 2.29) is essential as it provides a useful system of reference to regions of the head, but a detailed knowledge of the disarticulated units has little practical value.
Conventional descriptions are based on the views obtained from various directions with the skull resting on a flat surface. In most views the two distinct portions of the skull are immediately apparent: the caudal part encasing the brain and the rostral part supporting the face. The orbits, the fossae containing the eyeballs, are part of the face but lie at the boundary. In most domestic animals the facial part of the skull is larger than the neural part and is situated mainly in front of this. However, the ratio varies among species and also with breed, age, and individual conformation. The many particular differences make it impossible to provide even a general description of the skull that is valid for all species.
The Skull of the Dog
This initial account is of the skull of an adult dog of average (mesaticephalic) conformation, neither short-headed (brachycephalic) like a Pekingese nor long-headed (dolichocephalic) like a Borzoi. Some salient breed differences are mentioned later (p. 359).
» TABLE 2.1
Derivatives of the Pharyngeal Arches
In the dorsal view (Fig. 2.30), the ovoid cranium meets the bones of the face where the zygomatic processes (Fig. 2.30/4') of the frontal bones project laterally to form the dorsocaudal parts of the orbital walls. The caudal extremity of the cranium is marked by the external occipital protuberance in the midline, and its demarcation from the caudal (nuchal) surface is completed by the nuchal crests that extend laterally to each side. The median sagittal crest that extends forward from the occipital protuberance is most prominent in robust, well-muscled animals. All these features are easily palpated in life. The dorsal and lateral surfaces of each half of the cranium blend in a continuous and slightly roughened surface from which the temporalis muscle arises. Rostral to the zygomatic processes of the frontal bones the dorsal surface of the skull dips, sometimes quite markedly, before continuing as the straight and narrow dorsum of the nose. This ends at the wide nasal aperture, beyond which the bony skull is prolonged by pliant nasal cartilages.
The orbit is the most prominent feature of the lateral view (Fig. 2.31). Behind the orbit, the dorsolateral part of the braincase forms the wall of the temporal fossa (Fig. 2.31/16). The ventrolateral part is more complicated and presents the zygomatic arch and ear regions. The zygomatic arch (Fig. 2.31/15) springs free from the braincase and, bowing laterally, passes below the orbit to rejoin the facial part of the skull. It is formed by two bones, the squamous temporal and the zygomatic, which meet at an overlapping suture.
The ventral surface of the caudal part of this arch is shaped like a transverse gutter for articulation with the mandible; the articular area continues caudal to the rostral surface of the retroarticular process (Fig. 2.31/6). The large, smooth dome of the tympanic bulla (Fig. 2.31/9) (enclosing part of the cavity of the middle ear) and the rough mastoid process lie behind the retroarticular process. Three openings are present in this region of the skull: the retroarticular foramen emits a major vein draining the cranial cavity; the stylomastoid foramen gives passage to the facial nerve; and the external acoustic meatus is, in the fresh state, closed by a membrane (eardrum) that separates the canal of the external ear from the cavity of the middle ear. The paracondylar process (Fig. 2.31/11) is conspicuous at the caudal limit of the skull.The orbit is funnel shaped, and in the macerated state its walls are very incomplete. In life the orbital rim is completed by a ligament (Fig. 2.31/1) that connects the zygomatic process of the frontal bone to the zygomatic arch. Ventrally the orbital cavity is continuous with the pterygopalatine fossa (Fig. 2.31/4), but in the fresh state these regions are separated by the periorbita, a dense fascial sheet that completes the definition of the orbit. Two groups of foramina are visible in this region. The caudal group (Fig. 2.31/5) comprises the optic canal, orbital fissure, and rostral alar foramen. The optic opening, placed at the apex of the conical orbital cavity, is the portal of entry of the optic nerve. The more ventral orbital fissure transmits the nerves (ophthalmic, oculomotor, trochlear, and abducent) that supply ancillary structures of the eye and the external ophthalmic vein. Most ventrally the rostral alar foramen provides a common opening for the maxillary nerve, passing from the cranial cavity, and the maxillary artery, which traverses a canal (alar canal) in the sphenoid bone.
The rostral group of foramina comprises the maxillary, sphenopalatine, and caudal palatine foramina.
The maxillary foramen (Fig. 2.30/2') leads to the infraorbital canal, the sphenopalatine foramen to the nasal cavity, and the caudal palatine to the palatine canal, which emerges on the hard palate; each opening conveys like-named branches of the maxillary artery and nerve. More dorsally the rostral orbital wall contains the lacrimal fossa for the lacrimal sac (Fig. 2.30/3). An opening in the depth of the fossa leads to a passage that conveys the nasolacrimal (tear) duct to the nose.The infraorbital foramen (Fig. 2.30/2), the most prominent and easily palpable feature of the lateral aspect of the face, provides exit to the infraorbital nerve, which continues from the maxillary nerve through the infraorbital canal. Toward the alveolar margin the facial skeleton is molded over the roots of the teeth, most especially over the large root of the canine tooth.
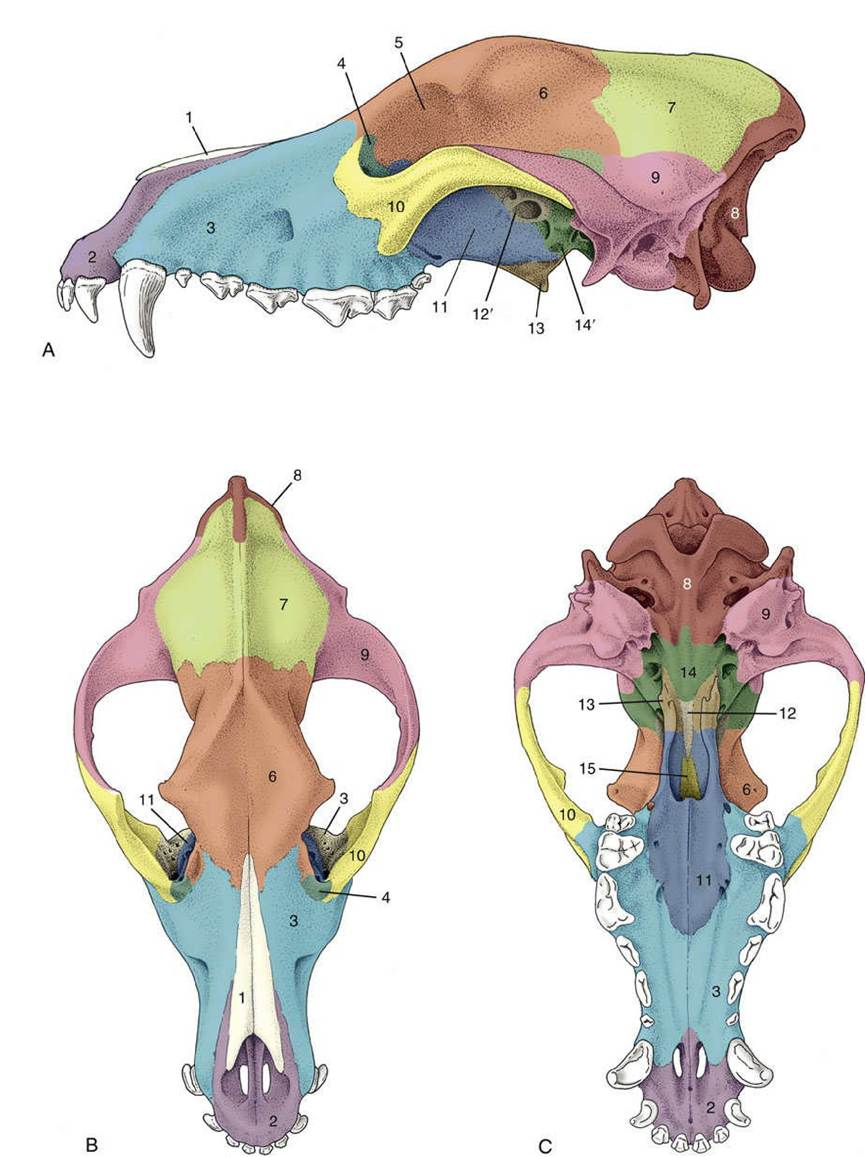
FIG. 2.29 (A) Lateral, (B) dorsal, and (C) ventral views of the canine skull to show the extents of the cranial bones. 1, Nasal bone; 2, incisive bone; 3, maxilla; 4, lacrimal bone; 5, orbit; 6, frontal bone; 7, parietal bone; 8, occipital bone; 9, temporal bone; 10, zygomatic bone; 11, palatine bone; 12, presphenoid; 12', wing of presphenoid; 13, pterygoid bone; 14, basisphenoid; 14, pterygoid process of basisphenoid; 15, vomer.
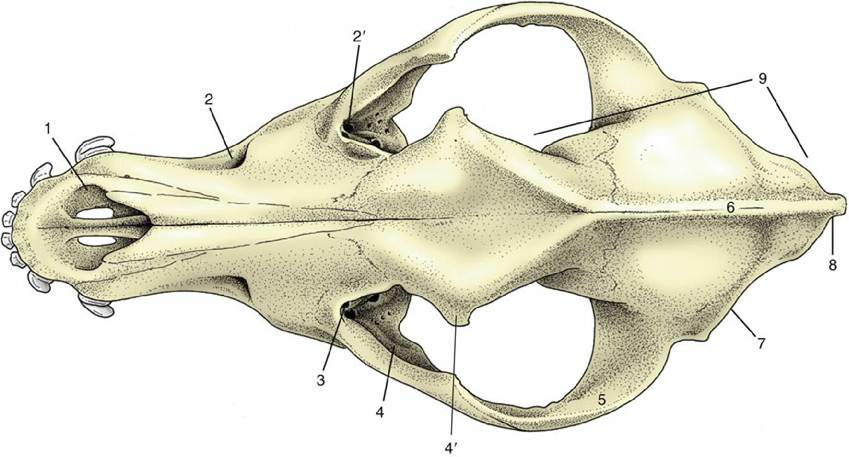
FIG. 2.30 Dorsal view of canine skull. 1, Nasal aperture; 2, infraorbital foramen; 2', maxillary foramen; 3, fossa for lacrimal sac; 4, orbit; 4', zygomatic process of frontal bone; 5, zygomatic arch; 6, external sagittal crest; 7, nuchal crest; 8, external occipital protuberance; 9, cranium.
In the ventral view (Fig. 2.32), three regions of the skull are distinct: the base of the cranium, the choanal region where the nasal cavities open into the pharynx, and the hard palate. The first shows at its caudal limit the ovoid, obliquely oriented occipital condyles that flank the foramen magnum (Fig.
2.32/12), through which the spinal cord connects with the brain. Rostral to this the median area is generally flat, although midway along its length, tubercles are present for the attachment of muscles that flex the head on the neck. The tympanic bulla and paracondylar process occupy much space to each side. The medial aspect of the bulla (Fig. 2.32/7) meets the occipital bone, and this fusion separates two openings that are confluent in some other species (e.g., horse; see Fig. 2.37) — namely, the more caudal jugular foramen and the more rostral foramen lacerum (Fig. 2.32/8 and 6). The glossopharyngeal, vagus, and accessory nerves emerge through the jugular foramen together with a large vein draining the interior of the cranium. Between the jugular foramen and the condyle is the hypoglossal canal, which transmits the hypoglossal nerve.Lateral to the foramen lacerum, small fissures exist for the exit of the chorda tympani (a branch of the facial nerve) and for the communication of the cartilaginous auditory tube with the cavity of the middle ear. Rostral to these is the prominent oval foramen (Fig. 2.32/4), through which the mandibular nerve emerges.
The openings (choanae) that lead from the nasal cavities to the nasopharynx are the main features of the middle part of the ventral aspect. The choanal region is bounded dorsally by the floor of the cranium and laterally by the thin plates of bone whose outer surfaces were earlier noted as forming the medial walls of the pterygopalatine fossae. The soft palate, which arises from the free margin of the hard palate, in life provides the floor of the space—essentially the first part of the nasopharynx —enclosed by these formations. The palate, which lies rostral to nasopharynx, is broad behind and narrower in front. It is margined by the alveoli or sockets in which the upper teeth are implanted. Toward its rostral extremity, it is perforated by the large bilateral palatine fissures. Several smaller foramina toward the caudal extremity of the palate are rostral openings of the palatine canal.
The nuchal surface (Fig. 2.31/13), broadly triangular, is limited dorsally by the external occipital protuberance and the nuchal crests. Its lower part presents the foramen magnum, the occipital condyles, and the paracondylar processes. The remainder of the surface is roughened for the attachment of dorsal muscles of the neck.
The apex of the skull is formed by the nasal aperture situated dorsal to the rostral extremities of the jaws that carry the incisor teeth.
The cavities of the skull are described with the respiratory system (Chapter 4), central nervous system (Chapter 8), and ear (Chapter 9).
The lower jaw or mandible comprises two parts (Fig. 2.33). In the dog these are firmly but not rigidly united by the connective tissues of the mandibular symphysis. Each half is divided between a body, or horizontal part, and a ramus, or vertical part. The body carries the alveoli of the lower teeth and is laterally compressed. Except at its rostral extremity, it diverges from its fellow to bound an intermandibular space. Toward its rostral extremity the lateral surface presents several mental foramina, one generally much larger than the rest; through these emerge the mental branches of the inferior alveolar nerve and vessels. The ramus (Fig. 2.33/2) is wider but less robust. Its dorsal extremity ends in the coronoid process, which projects into the temporal fossa and gives attachment to the temporalis muscle, and the lower and more caudal condylar process (Fig. 2.33/3), which carries an articular head shaped like a portion of a truncated cone. The lower part of the caudal margin of the ramus carries the projecting angular process that gives attachment to the masseter and medial pterygoid muscles. The lateral surface has a roughened depression where the masseter inserts. The medial surface gives insertion to the pterygoid muscles and also presents the large mandibular foramen (Fig. 2.33/7) for entry of the inferior alveolar vessels and nerve.
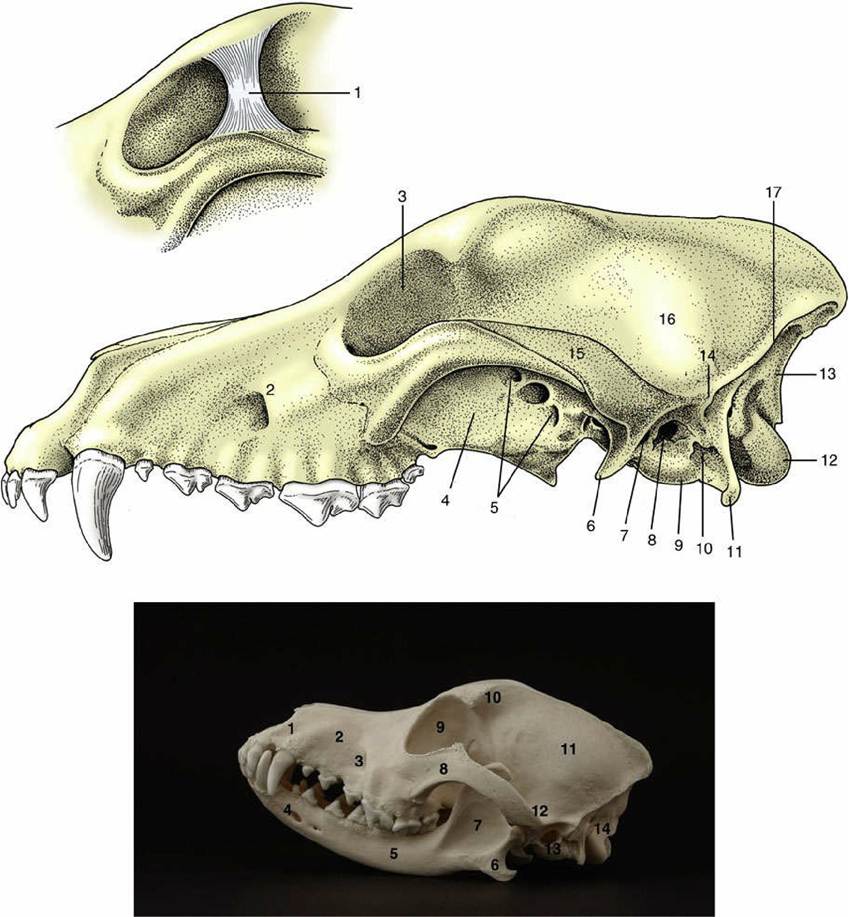
FIG. 2.31 Lateral view of canine skull. 1, Orbital ligament (inset); 2, infraorbital foramen; 3, orbit; 4, pterygopalatine fossa; 5, optic canal, orbital fissure, and rostral alar foramen; 6, retroarticular process; 7, retroarticular foramen; 8, external acoustic meatus; 9, tympanic bulla; 10, stylomastoid foramen; 11, paracondylar process; 12, occipital condyle; 13, nuchal surface; 14, mastoid process; 15, zygomatic arch; 16, temporal fossa; 17, nuchal crest.
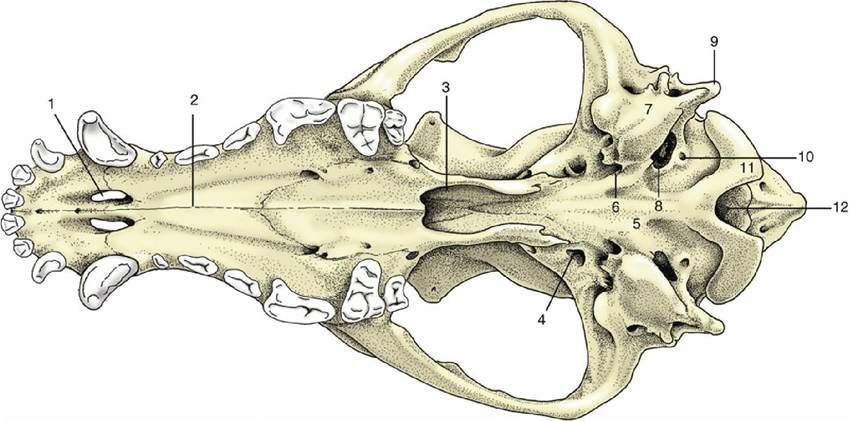
FIG. 2.32 Ventral view of canine skull. 1, Palatine fissure; 2, hard palate; 3, choanal region; 4, oval foramen; 5, base of cranium; 6, foramen lacerum; 7, tympanic bulla; 8, jugular foramen; 9, paracondylar process; 10, hypoglossal canal; 11, occipital condyle; 12, foramen magnum.
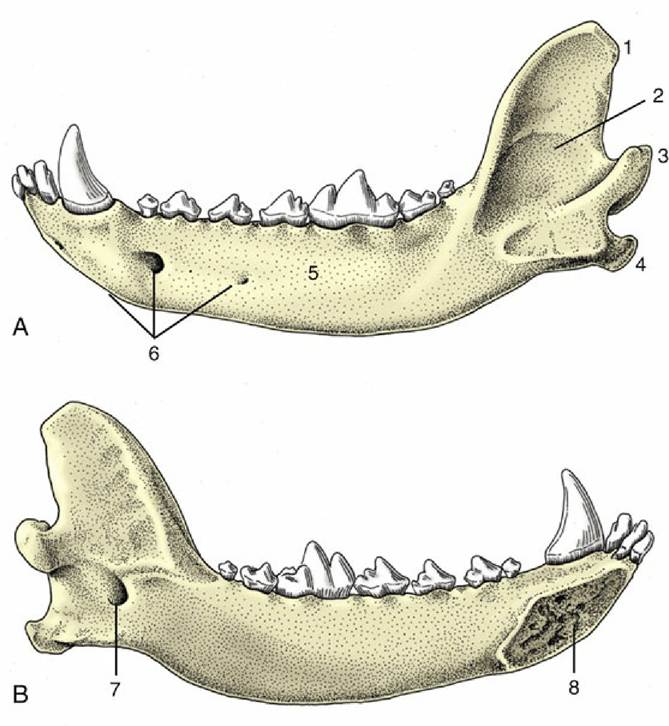
FIG. 2.33 (A) Lateral and (B) medial views of the left half of the canine mandible. 1, Coronoid process; 2, vertical part (ramus); 3, condylar process; 4, angular process; 5, horizontal part (body); 6, mental foramina; 7, mandibular foramen; 8, symphysial surface.
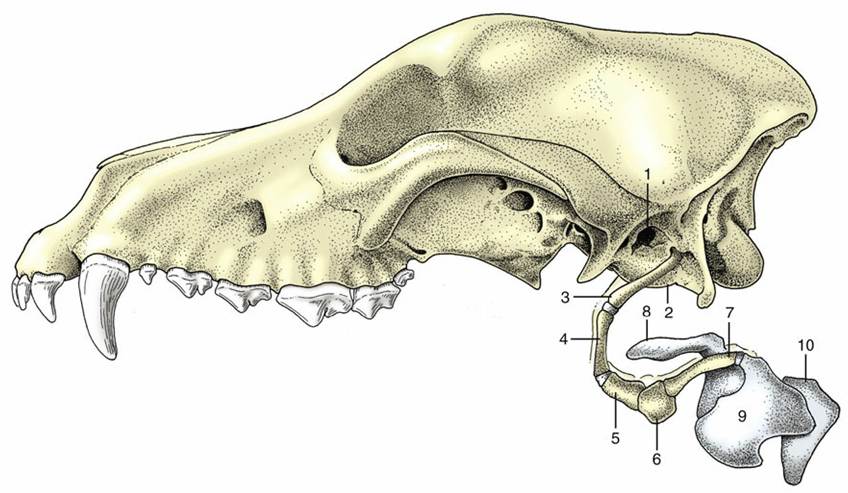
FIG. 2.34 Hyoid apparatus and larynx suspended from the temporal region of a canine skull. 1, External acoustic meatus; 2, tympanic bulla; 3, stylohyoid; 4, epihyoid; 5, ceratohyoid; 6, basihyoid; 7, thyrohyoid;
8, epiglottic cartilage; 9, thyroid cartilage; 10, cricoid cartilage.
The hyoid apparatus is a series of bony rods jointed together and forming a means of suspending the tongue and larynx from the skull. Fig. 2.34 shows the names of the several parts, their arrangement, and the attachment of the apparatus as a whole to the temporal region of the skull. The transversely placed basihyoid may be palpated within the intermandibular space; other parts are palpable—indeed their positions are visible—when the walls of the pharynx are inspected through the mouth.
Some Comparative Features of the Skull
When equipped with the mandible the skull of the cat (Fig. 2.35) appears globular. Several features combine to create this conformation:
• the rounded cranial capsule, surmounted by a short, often weak sagittal crest, and corresponding closely to the contours of the brain
• the very salient convex zygomatic arches
• the relative shortness of the face, which may account for as little as 20% of the total length
The orbital region is distinctive. The orbits are large, face more directly forward than in the dog, and have more complete bony margins. The frontal process of the zygomatic bone and the zygomatic process of the frontal bone leave only a small gap in the ovoid margin to be closed by the orbital ligament. The zygomatic arch is surprisingly strong where it contributes to the orbital rim. The infraorbital foramen is placed close to the rostroventral part of the orbit, where it may be palpated.
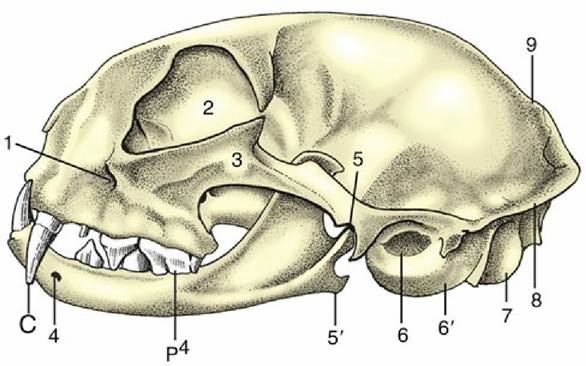
FIG. 2.35 Feline skull with mandible. 1, Infraorbital foramen; 2, orbit; 3, zygomatic arch; 4, mental foramen; 5, temporomandibular joint; 5', angular process of mandible; 6, external acoustic meatus; 6', tympanic bulla; 7, occipital condyle; 8, nuchal crest; 9, sagittal crest; C, canine tooth; p4, upper fourth premolar.
On the ventral aspect, the hard palate is short and wide, and it carries alveoli for only four cheek teeth. That for the largest of these teeth (P4) is located dangerously close to the orbit, which may become involved in a spreading alveolar abscess. Caudally, the deep gutter of the temporomandibular articulation is bounded by a prominent retroarticular process. The very large tympanic bulla may be palpated between the caudal part of the zygomatic arch and the wing of the atlas.
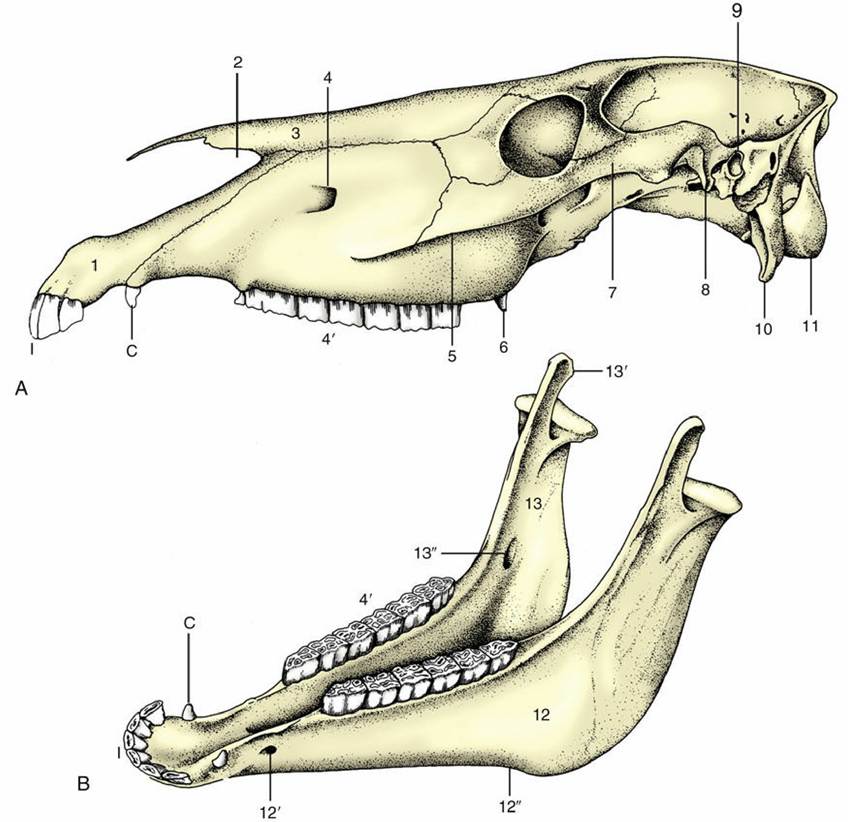
FIG. 2.36 (A) Equine skull and (B) equine mandible. 1, Incisive bone; 2, nasoincisive notch; 3, nasal bone; 4, infraorbital foramen; 4', cheek teeth; 5, facial crest; 6, hamulus of pterygoid bone; 7, zygomatic arch; 8, retroarticular process; 9, external acoustic meatus; 10, paracondylar process; 11, occipital condyle; 12, horizontal part (body) of mandible; 12', mental foramen; 12", vascular notch; 13, vertical part (ramus) of mandible; 13', coronoid process; 13", mandibular foramen; I, incisors; C, canine tooth (present only in the male).
As in the dog, the halves of the mandible do not fuse, even in old age, and a small degree of movement is allowed at the mandibular symphysis. Each half carries sockets for only three cheek teeth.
Breed differences are more pronounced than sometimes supposed. The skulls of Siamese and similar cats have much longer faces, which often blend smoothly with the cranium without any break (stop) in the dorsal contour. In contrasting types—for example, the Persian—the face is short and shallow, and the stop is prominent.
The equine skull (Fig. 2.36) is characterized by a relatively long face, a feature that develops further with increasing size; it is therefore more pronounced in mature than in juvenile animals and in large than in small breeds. The cranium is relatively narrow and generally not unlike that of the dog. The external sagittal crest is weaker. The forehead is wide between the origins of the zygomatic processes of the frontal bones, which bend ventrally to join the zygomatic arches.
The zygomatic arch (Fig. 2.36/7) is conspicuously strong, even without the extra support it obtains from the zygomatic process connecting it with the frontal bone taken into account. It is not bowed laterally to any extent and carries a rather complicated articular surface on its caudoventral aspect; this comprises a rostral tuber, an intermediate fossa, and a salient retroarticular process (Fig. 2.36/8). The orbit faces almost laterally and has a complete bony rim. A large maxillary tuberosity appears to continue the alveolar process directly. The zygomatic arch is continued rostrally, beyond the orbit, as a prominent ridge on the lateral surface of the face. This ridge, the facial crest (Fig. 2.36/5), runs parallel to the dorsal contour of the nose and ends above a septum between the alveoli of the third and fourth cheek teeth in the adult.
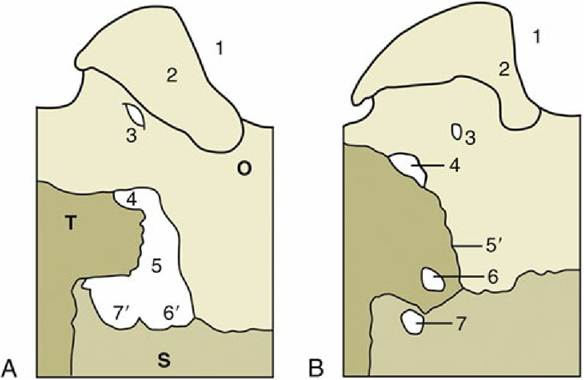
FIG. 2.37 Left Caudolateral parts of the base of the (A) equine and (B) canine cranium, showing portions of the occipital (O), sphenoid (S), and temporal (T) bones; ventral view (schematic). 1, Foramen magnum;
2, occipital condyle; 3, hypoglossal canal; 4, jugular foramen; 5, foramen lacerum; 5', petro-occipital suture; 6, foramen lacerum; 6', carotid notches; 7, oval foramen; 7', oval notch.
A deep (nasoincisive) notch separates the pointed nasal bone from the incisive bone (Fig. 2.36/1— 3). This notch and the rostral end of the facial crest are both very easily identified landmarks; they are used as guides to the position of the infraorbital foramen, which lies a little caudal to the middle of the connecting line (Fig. 2.36/4).
The features visible on the ventral view lie more or less on one level. The caudal part of this surface is distinguished by the large and very salient paracondylar processes (Fig. 2.36/10) and the jagged outlines of the large openings to each side of the occipital bone. Each opening results from the failure of the temporal bone to reach the lateral margin of the occipital bone, which permits the confluence of several foramina that are distinct in the dog. The caudal part is the equivalent of the jugular foramen; the cranial part (foramen lacerum) combines the oval and carotid foramina (Fig. 2.37/7 and 6). In life the greater part of the large opening is occluded by membrane that leaves barely sufficient passage for the various nerves and vessels. The tympanic bulla is not prominent, but the styloid (for the hyoid apparatus) and muscular processes of the temporal bone are well developed.
The choanae lie almost in the plane of the hard palate. The vertical plate of bone that separates the choanal from the pterygopalatine region carries a prominent hamular process (Fig. 2.36/6). The greater part of the margin of flat and unremarkable palate is occupied by the alveoli of the incisor and cheek teeth. A well-marked external occipital protuberance is present on the nuchal surface, midway between the nuchal crest and the dorsal margin of the foramen magnum.
The mandible has the following features:
• It is massive, and its right and left halves diverge at a relatively small angle (Fig. 2.36B).
• Its symphysis becomes obliterated quite early, usually about 2 years after birth.
• Its lower margin carries a prominent vascular notch where the facial vessels wind onto the face (Fig. 2.36/12"").
• Its ramus is high, the coronoid process projects far into the temporal fossa, and the articular process carries the ovoid articular surface well above the occlusal plane of the cheek teeth.
The parts of the hyoid apparatus (see Fig. 4.8) are of different proportions to their counterparts in the dog and are laterally compressed. A substantial lingual process projects from the basihyoid into the root of the tongue.
The bovine skull (Fig. 2.38) is relatively short and wide with a general pyramidal form. Cornual (horn) processes project from the frontal bones of horned breeds where the dorsal, lateral, and nuchal surfaces meet; their size and direction vary greatly with breed, age, and sex. The very wide and flat frontal region is bounded by a prominent temporal line that overhangs the deep temporal fossa and confines it to the lateral aspect of the skull.
The principal features of the lateral aspect are the confinement of the temporal fossa and the elevation of the orbital rim above its surroundings. The rim is complete and is formed by the meeting of processes from the zygomatic and frontal bones in its caudal part. There is no facial crest, only a discrete facial tuberosity from which the rostral part of the masseter arises. The infraorbital foramen is directly above the first cheek tooth, rather low toward the palate.
The ventral surface is very uneven, and the cranial base is located in a considerably more dorsal plane than the palate. The temporal and occipital bones are separated by a narrow fissure, which is an arrangement intermediate between the suture of the dog and the wide opening of the horse and pig. The tympanic bulla is prominent and laterally compressed. The choanae are separated by the caudal prolongation of the ventral part of the nasal septum and are enclosed laterally by very extensive plates of bone. The palate, long and narrow, is bounded by high alveolar processes. Of course, no alveoli are present for incisor or canine teeth, which are lacking in the upper jaws of ruminants.
The mandibular symphysis ossifies late, if at all, in ruminants. In general, the mandible is weaker than that of the horse, which is a feature very apparent in the body of the bone with its gently convex ventral border. The coronoid process is high and caudally inflected. The articular surface is concave and widened laterally.
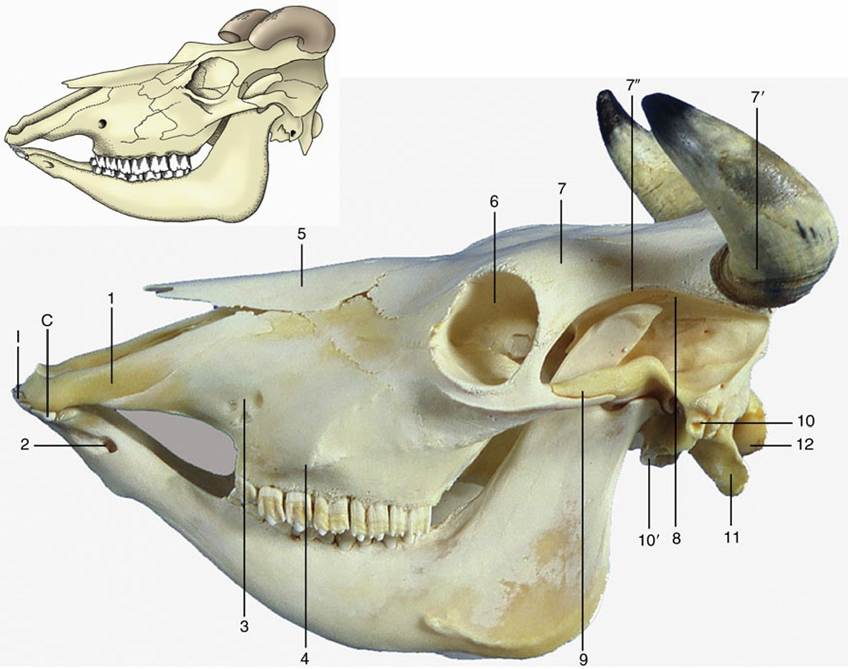
FIG. 2.38 Bovine skull with mandible. 1, Incisive bone; 2, mental foramen; 3, infraorbital foramen; 4, facial tuberosity; 5, nasal bone; 6, orbit; 7, frontal bone; 7', horn surrounding cornual process of frontal bone; 7", temporal line; 8, temporal fossa; 9, zygomatic arch; 10, external acoustic meatus; 10', tympanic bulla; 11, paracondylar process; 12, occipital condyle; I, incisors; C, canine tooth, incorporated in the row of incisors.
The few remarks necessary regarding the skulls of the small ruminants and the pig are found on pages 634 and 739, respectively.
The Joints of the Head
The articulations between the skull and mandible (temporomandibular joints) and between the halves of the mandible (mandibular symphysis) are appropriately considered in the following chapter (p. 104) because the teeth, the muscles of mastication, and the joints form a single functional complex.
The Muscles of the Head and Ventral Part of the Neck
The principal groups into which the muscles of the head may be divided are listed in Table 2.2, which draws attention to the correspondence between embryologic origin, innervation, and function. The functional associations are so well defined and specific that it is more convenient to include most of them in other chapters, with related organs.
The first four groups take origin in the unsplit mesoderm, which covers the lateral and ventral walls of the pharynx and condenses to form the cores of the pharyngeal arches.
In lower vertebrates the muscles equivalent to the last two groups in Table 2.2 are known to develop from somites that appear to each side of the hindbrain, some rostral to the otocyst, the primordium of the inner ear, and the others caudal to it. A similar origin may be assumed in mammals, although the evidence for the formation of these somites is unconvincing at least. They are of course somatic muscles with the appropriate type of innervation.
The Trigeminal Musculature
The muscles of mastication constitute the greater part of the musculature supplied by the mandibular division of the trigeminal nerve, the motor nerve to the first pharyngeal arch. They are described in the chapter on the digestive system (p. 105). The same chapter deals with the digastricus — a composite muscle to which the mandibular field makes a contribution; the mylohyoideus which slings the tongue between the lower jaws; and one of the muscles (tensor veli palatini) of the soft palate (p. 109). The tensor tympani is considered with the middle ear (p. 333).
» TABLE 2.2
Source and Innervation of the Principal Muscle Groups of the Head
| Muscle Group | Source | Innervation |
| Masticatory musculature | First pharyngeal arch | Mandibular division of trigeminal nerve (V3) |
| Mimetic musculature | Second pharyngeal arch | Facial nerve (VII) |
| Fharyngeal and palatine musculature | Third and fourth pharyngeal arches | Glossopharyngeal (IX) and vagus (X) nerves |
| Laryngeal musculature | Sixth pharyngeal arch | Vagus nerve (X) |
| External ocular musculature | Hypothetical preotic somites | Oculomotor (III), trochlear (IV), and abducent (VI) nerves |
| Lingual musculature | Hypothetical postotic somites | Hypoglossal nerve (XII) |
The Facial Musculature
The musculature supplied by the facial nerve, the nerve of the second pharyngeal arch, is resolvable into two divisions. The superficial division comprises the cutaneous muscle of the head and neck in addition to many small units that control the posture of the lips, cheeks, nostrils, eyelids, and external ears. The deep division is rather scattered but includes some muscles associated with the hyoid apparatus, a contribution to the digastricus (p. 109), and the stapedius (p. 334) of the middle ear.
The Superficial Division
The muscles of the superficial division are conjectured to have their source in an ancestral deep sphincter muscle of the neck, which may be envisaged as arranged in three incomplete overlapping layers. The outermost layer, consisting of transversely disposed fascicles, is reduced to insignificance or is entirely lacking in domestic mammals. A remnant (sphincter colli) survives in the dog. A more substantial portion of the middle layer commonly persists as platysma organized as a sheet of longitudinally disposed fibers covering the ventral part of the face and extending onto the neck, even reaching the nape in the dog. Detached slips are believed to provide the small muscles that attach to the caudal aspect of the external ear.
The third and deepest layer is also transverse and is believed to be the origin of the many discrete muscles of the mammalian face. These are extremely variable among species, but very few require detailed notice. Because of their effect on the appearance of the face, they are collectively known as the muscles of facial expression or mimetic musculature.
The principal muscles of the lips and cheeks are the buccinator, orbicularis oris, caninus, levator nasolabialis, levator labii superioris, and depressor labii inferioris (Figs. 2.39 and 11.6). The buccinator (Fig. 2.39/4) passes between the margins of the upper and lower jaws and is partly covered by the masseter. It forms the basis of the cheek and opposes the tongue to prevent food from collecting in the vestibule by returning it to the central cavity of the mouth. The buccal salivary glands are scattered among its fascicles, and discharge of their secretion into the mouth may be assisted by contraction of the muscle. The orbicularis oris (Fig. 2.39/1) surrounds the mouth opening, where it is closely attached to the skin and mucosa of the lips. It closes the opening of the mouth by pursing the lips and is important in sucking. The caninus (Fig. 2.39/2) arises ventral to the infraorbital foramen and radiates into the wing of the nostril and the upper lip. It dilates the nostril and elevates the corner of the mouth in the snarling gesture, especially in the dog. The levator nasolabialis (Fig. 2.39/5) arises over the dorsum of the nose and inserts partly on the wing of the nostril and partly into the lateral part of the upper lip. It is able to dilate the nostril and to elevate and retract the upper lip. The medial part of the upper lip is elevated by the separate levator labii superioris (Fig. 2.39/6). This muscle arises on the lateral aspect of the face and runs dorsorostrally to form with its fellow a common tendon that descends into the lip between the nostrils. A special depressor labii inferioris is present in the lower lip of certain species (excluding the dog and cat). It appears to be a detachment from the buccinator muscle. Other muscles associated with the lips and nostrils do not merit specific mention, although some are identified in various illustrations.
The muscles of the eyelids include one, the levator palpebrae superioris, that is clearly foreign to the facial group because it arises within the orbit and is supplied by the oculomotor nerve (see p. 328). The muscles of the lids that are supplied by the facial nerve include a sphincter—the orbicularis oculi (Fig. 2.39/7)—that surrounds the palpebral fissure, the opening between the lids. It is anchored at the medial and lateral commissures and therefore narrows the opening to a horizontal slit when it contracts. Other muscles are present to raise the upper (levator anguli oculi) lid and to depress the lower (malaris) lid, enlarging the eye opening.
The muscles of the external ear are especially numerous but of little account individually. A caudal group has already been mentioned. Others converge on the auricle—the skin-covered cartilaginous ear "trumpet" — from medial, rostral, and lateral directions; they lie between the skin and the temporalis muscle and skull and form a thin, incomplete sheet that includes a (scutiform) cartilage plate. The scattered origins and precisely located insertions provide for displacement and rotation of the ear in all directions. One, the parotidoauricularis, is of somewhat greater importance because it is encountered in the operation for drainage of infections of the external ear of the dog (p. 381). As its name suggests, it arises from the fascia over the parotid gland and approaches the auricle from the ventrolateral direction.
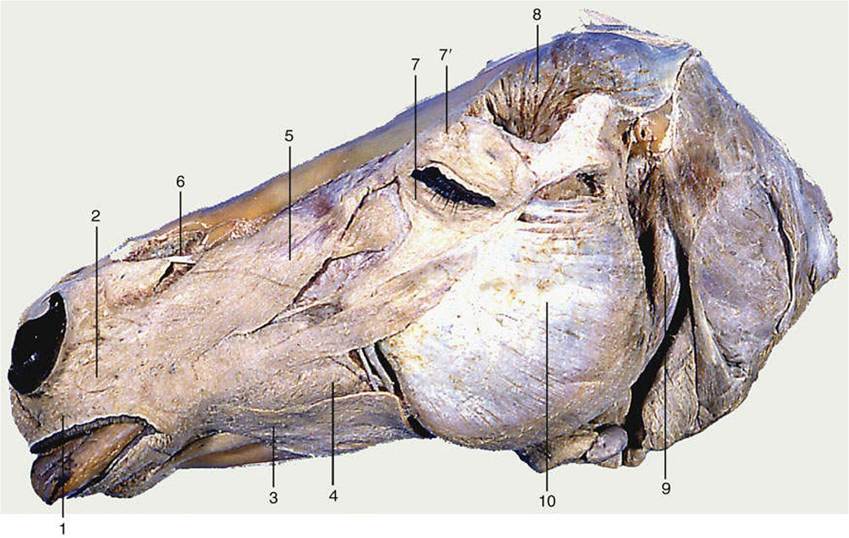
FIG. 2.39 Superficial muscles of the equine head. The cutaneous muscle has been removed. 1, Orbicularis oris; 2, caninus; 3, depressor labii inferioris; 4, buccinator; 5, levator nasolabialis; 6, levator labii superioris; 7, orbicularis oculi; 7', levator anguli oculi medialis; 8, temporalis; 9, occipitomandibular part of digastricus; 10, masseter.
Besides the individual functions mentioned or implied in the preceding paragraphs, these muscles have a collective function in communication, mainly within the species but also between species. Human observers can intuitively, or as the result of experience, interpret many obvious facial gestures of animals: one need only recall the hangdog expression of submission, the evident threat conveyed by snarling or laying back the ears, or the quizzical look a dog may adopt.
Paralysis of these muscles is not uncommon after damage to the facial nerve. Because different groups are supplied by branches of the nerve that arise at different levels, the particular pattern of distortions can be a valuable pointer to the location of the nerve lesion (p. 305).
The Deep Division
The muscles attaching to the hyoid apparatus are a rather heterogeneous assemblage. Although certain small units supplied by the facial nerve elevate the hyoid, to draw the tongue backward to aid in swallowing, the muscles do not appear to merit description. The digastricus, in part derived from the facial musculature, is described on page 105; the stapedius of the middle ear is described on page 333.
The Muscles of the Pharynx and Soft Palate
The muscles of the pharynx and soft palate are considered beginning on page 107.
The Muscles of the Larynx
The laryngeal muscles are considered beginning on page 144.
The External Muscles of the Eyeball
The external muscles of the eyeball are considered beginning on page 327.
The Muscles of the Tongue
The tongue muscles are considered beginning on page 95.
The Muscles of the Ventral Part of the Neck
The neck connects the head with the trunk and is usually distinguished by its relatively slender construction—although this is hardly true of the pig. The neck has a generally cylindrical form in the dog and cat but is quite obviously compressed from side to side in the larger animals, in which it deepens considerably toward its junction with the thorax (Fig. 2.40). The core structures of the neck—the cervical vertebrae and the muscles closely applied to them—were described with the trunk (p. 42). Certain superficial muscles are considered under the heading of "Girdle Muscles of the Forelimb" (p. 75). The present section is therefore concerned only with the ventral part of the neck, a region of considerable clinical importance on account of the numerous visceral, vascular, and nervous structures that traverse it en route between the head and thorax.
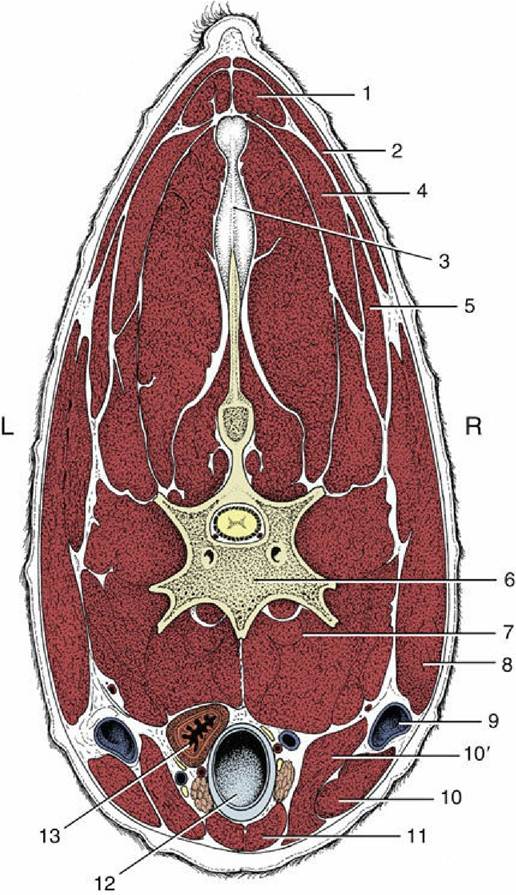
FIG. 2.40 Transverse section of the bovine neck. 1, Rhomboideus; 2, trapezius; 3, nuchal ligament; 4, splenius; 5, omotransversarius; 6, vertebra; 7, longus colli; 8, brachiocephalicus; 9, external jugular vein in jugular groove; 10 and10', sternocephalicus, mandibular and mastoid parts; 11, combined sternohyoideus and sternothyroideus; 12, trachea; 13, esophagus (ventral to it, nerves, blood vessels, and thymus); L, left; R, right.
These structures, with the important exception of the external jugular veins (Fig. 2.40/9), occupy a central visceral space. The roof of this space is provided by the muscles immediately ventral to the vertebrae—namely, the longus colli, longus capitis, rectus capitis ventralis, and scalenus (p. 44). The side and ventral walls blend together and are provided by thinner muscles disposed with a sagittal course and joined by stout fasciae.
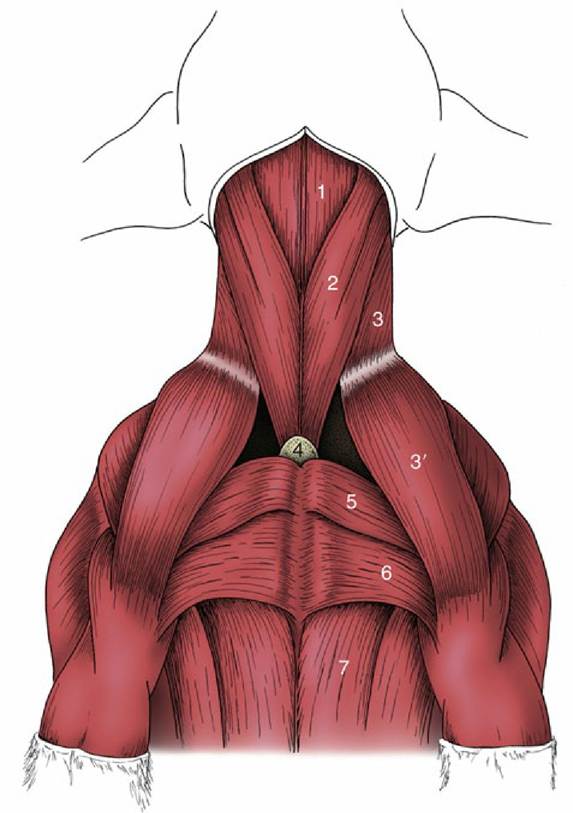
FIG. 2.41 Ventral muscles of the canine neck and thorax. 1, Combined sternohyoideus and sternothyroideus; 2, sternocephalicus; 3 and 3', brachiocephalicus: cleidocervicalis and cleidobrachialis; 4, manubrium of sternum; 5, pectoralis descendens; 6, pectoralis transversus; 7, pectoralis profundus.
The cervical part of the cutaneous muscle (m. cutaneous colli) is unimportant in the dog and cat. It is much better developed in the ungulates, in which it radiates from a stout origin on the manubrium of the sternum; it thins as it passes cranially and laterally and eventually fades away. In the horse, the cutaneous muscle provides a relatively thick cover to the caudal third or so of the jugular groove.
The straplike sternocephalicus (Fig. 2.41/2) is the most ventral of the other muscles. It also arises from the manubrium, ascends the neck, and diverges laterally toward its insertion, which varies among species but includes one or the other (or both) of the angle of the mandible and the mastoid process of the skull. The divergence of the right and left muscles leaves a very thin layer of the deeper muscle, enabling palpation of the trachea through the skin. The sternocephalicus is supplied by the ventral branch of the accessory nerve. Unilateral contraction draws the head and neck to that side. Bilateral contraction flexes the head and neck ventrally. In species with a mandibular insertion the sternocephalicus may assist in opening the mouth.
The sternocephalicus forms the ventral border of the jugular groove that houses the external jugular vein (Fig. 2.42). The dorsal border of the groove is furnished by the brachiocephalicus, described more fully elsewhere (p. 76). The groove is often visible in life, particularly toward the upper part of the neck.
The deeper infrahyoid group of muscles provides an incomplete cover to the lateral and ventral aspects of the trachea and inserts, directly or indirectly, on the hyoid apparatus, which they all stabilize and retract toward the thorax during swallowing. The obvious members of the group are the sternothyroideus, sternohyoideus, and omohyoideus; the thyrohyoideus on the lateral aspect of the larynx may be regarded as a detached member. The nerve supply is mainly, although possibly not entirely, from the first and second cervical nerves.
The sternothyroideus and sternohyoideus are very thin ribbonlike muscles that take a common origin from the manubrium of the sternum. The caudal parts of the right and left muscles are not always distinctly divided, and in the middle of the neck they may have a common intermediate tendon from which three or four slips diverge cranially. The sternothyroideus inclines laterally to terminate on the lateral aspect of the thyroid cartilage. The sternohyoideus, not always separable from its fellow, passes beside the midline to insert on the basihyoid.
The omohyoideus, lacking in carnivores, is also thin and straplike. Its absence is compensated by the relative enlargement of the other muscles. In the horse it arises from the subscapular fascia and in the ruminants from the deep fascia of the neck; thereafter it edges medially to join the lateral margin of the sternohyoideus beside which it inserts. In the horse the omohyoideus provides a floor to the caudal part of the jugular groove, separating the vein from the structures within the visceral space.