The Small-Intestinal Mucosa Has a Large Surface Area and Epithelial Cells with "Leaky" Junctions Between Them
Contact between the small-intestinal mucosa and the luminal contents is facilitated by an extensive intestinal surface area. Three levels of surface convolutions serve to expand the surface area of the small intestine (see Figure 27-2).
First, large folds of mucosa known as plicae circulares add to the intestinal surface area of some animals, but these are not present in all species. Second, the mucosal surface is covered with fingerlike epithelial projections known as villi. These structures are present in all species and increase the intestinal surface area by 10-fold to 14-fold compared with a fiat surface of equal size. Third, the villi themselves are covered with a brushlike surface membrane known as the brush border. I he brush border is composed of Submicroscopic microvilli that further enlarge the surface area (Figure 30-2). At the base of the villi are glandlike structures known as crypts of Lieberkuhn (Figure 30-3). The villi and crypts are covered with a continuous layer of cellular epithelium.The epithelial cells covering the villi and crypts are called enterocytes. Each enterocyte has two distinct types of cell membranes (Figure 30-4). The cell surface facing the lumen is called the apex and is covered by the apical membrane. The apical membrane contains the microvilli. Under the light microscope, the microvilli give the cell surface its brushlike appearance (brush border, synonymous with apical membrane). Covering the apical membrane, and encasing the microvilli, is a jellylike layer of glycoprotein known as the glycocalyx. Important digestive enzymes and other proteins are attached to the microvilli and project into the glycocalyx. Under the intense magnification of the electron microscope,

FIGURE 30-2 Electron micrograph of the microvilli of the intestinal brush border.
The brush border is composed ofthe apical membranes of e∩terocytes. Note the indistinct array of molecular material (S) that radiates away from the microvilli (V)'t membranous-phase digestion occurs within this array of molecular material, which includes the membrane-bound digestive enzymes. (From Johnson LR, Christensen J, Jacobsen ED, et al, editors: Physiology ofthe gastrointestinal tract, ed 2, NewYork, 1987, Raven Press.)these enzyme molecules, as well as other proteins, give the glycocalyx a fuzzy appearance (see Figure 30-2). The apical membrane is a complex and unusual cellular membrane with high protein content.
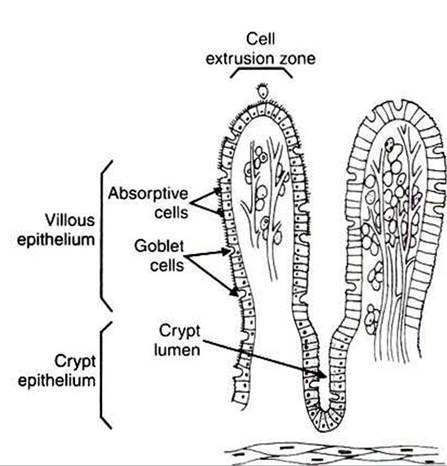
FIGURE 30-3 The one-cell-thick layer of intestinal epithelium is continuous over the villi and the crypts of Lieberkuhn.
The remaining portion of the enterocyte plasma membrane, that part not facing the gut lumen, is called the basolateral membrane, referring to the base and sides of the cell. This membrane is not especially unusual and indeed has many similarities to cell membranes of other tissues. Although the basolateral membrane is not in direct contact with ingesta in the gut lumen, it serves an important role in intestinal absorption; nutrients that are absorbed into the enterocytes through the apical membrane must exit the cell through the basolateral membrane before gaining access to the bloodstream.
'Γhe attachments between adjacent enterocytes are called tight junctions. These connections serve a special function in the process of digestion and absorption. The tight junctions form a narrow band Ofattachment between adjacent enterocytes. Γhe band is near the apical end of the cells and divides the apical membrane from the basolateral membrane. The junctions may be called “tight," but from a molecular standpoint, they are rather loose. This is especially true in the duodenum and jejunum, where the tight junctions are loose enough to allow the free passage of water and small electrolytes.
Recent evidence indicates that the relative impermeability, or “tightness," of the light junctions is not constant and can be altered by neurohumoral regulatory substances in the gut. These selective changes in permeability can affect the rates of water and ion movement across the gastrointestinal (GI) epithelium, depending on the physiological needs for secretion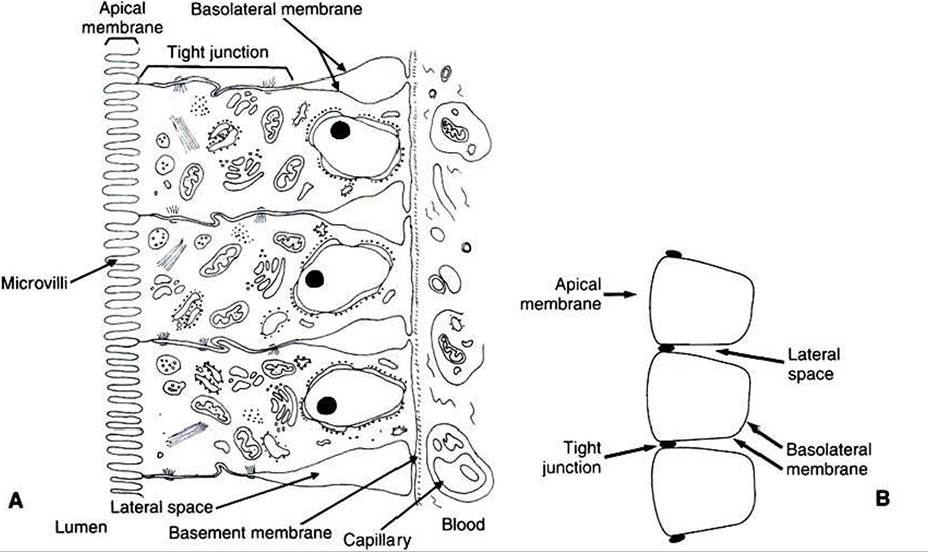
FIGURE 30-4 Understanding the anatomical relationships of the enterocytes, tight junctions, apical membrane, basolateral membrane, and lateral spaces is critical to an understanding of the physiology of intestinal absorption. A, Anatomic illustration of the intestinal epithelium. B, Stylized sketch of the epithelium, including a capillary containing formed elements of blood. It is important to understand the relationship between part A and part B of this diagram.
or absorption. However» the tight junctions are never permeable enough to permit the passage of organic molecules.
The narrow band of tight junctions leaves the majority of the basolateral membrane unattached to its neighboring membrane on the adjacent enterocyte. This arrangement creates a potential space between enterocytes. This area between the lateral surfaces of the enterocytes is called the lateral space. The lateral spaces are normally distended and filled with extracellular fluid (ECF). At the end of the lateral spaces nearest the apical membrane» the ECF is separated from the fluid of the intestinal lumen only by the tight junctions. Al the opposite end of the lateral spaces, the ECF is separated from the blood only by the basement membrane of the intestinal capillaries. Both the tight junctions and the capillary endothelium are permeable barriers that allow the free passage of water and small molecules. Thus, there is relatively free flow of water and most electrolytes among the fluid in the lumen of the intestine, the ECF in the lateral spaces, and the blood.