There Is GreaterVariety in the Specifics of Neuron-to-Neuron Synaptic TransmissionThan in Transmission at the Neuromuscular Junction
As mentioned earlier, some noteworthy differences exist between synaptic transmission at the neuromuscular junction and neuron-to-neuron synaptic transmission. Although acetylcholine is the neurotransmitter responsible for the primary postsynaptic effect at the neuromuscular junction, a variety of neurotransmitters, in addition to acetylcholine, can be used to produce the principal postsynaptic effect at neuron- to-neuron synapses (Box 5-1).
Eurthermore, not all of these transmitters are released from InorphoIogicallv distinct active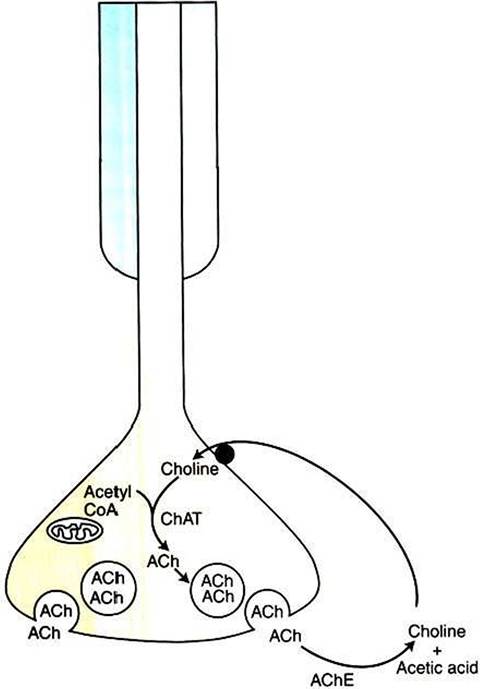
FIGURE 5-3 Synaptic inactivation of acetylcholine (ACh). Released ACh is enzymatically broken down into choline and acetic acid by synaptic acetylcholinesterase.The choline is actively transported back into the terminal and can be reused in ACh synthesis. ChAT, Choline acetyltransferase.
Box 5-1
Members of the Major Neurotransmitter Classes
Amino acids
Glutamate
Glycine
Y-Aminobutyric acid (GABA)
Amines
Acetylcholine
Serotonin
Histamine
Catecholamines
Dopamine
Norepinephrine
Epinephrine
Peptides[*]
Substance P
Vasopressin
Somatostatin
Opioids
Leu-enkephalin
Met-enkephalin
P-Endorphin
Nontraditional
Gases
Nitric oxide
Carbon monoxide
Purines
Adenosine
Adenosine triphosphate (ATP)
zones, although their release still appears to depend on Ca2+ influx. In such cases, release from the terminal may not always occur directly at the synaptic cleft, resulting in a wider postsynaptic distribution of transmitter.
The postsynaptic membrane of a neuron-to-neuron synapse can be the soma, dendrites, or even the terminals of the postsynaptic neuron, and junctional folds are not seen at these synapses. However, the dendritic postsynaptic membrane often possesses small protrusions called dendritic spines (see Chapter 4).
As with the junctional folds on muscle cells, these spines increase the surface area of the postsynaptic membrane and, because of their narrow necks, are also thought to provide a means for biochemical isolation between nearby synapses. Further, spines can change size and shape over an animals lifetime, modulating the functional effectiveness of the synapse. Whereas transmitter release at the neuromuscular junction always produces postsynaptic excitation (membrane depolarization), release at synapses between neurons can produce excitation or inhibition (membrane hyperpolarization). However, synapses on dendritic spines are almost always excitatory.At the neuromuscular junction, the postsynaptic receptor is almost exclusively the nicotinic acetylcholine receptor, a ligand-gated ion channel. At synapses between neurons, a much greater variety of receptors is available. These may differ from the nicotinic acetylcholine receptor with respect not only to the binding transmitter, but to the receptor mechanism as well (see Chapter 1). Also, several different types of neurotransmitter receptor are often found on a single neuron.
When transmitters other than acetylcholine are employed at neuron-to-neuron synapses, depending on the transmitter, the termination of action of that transmitter may be accomplished by (1) transporter-mediated reuptake of the transmitter itself into the terminal of release or (2) a less specific and somewhat slower form of enzymatic degradation than with acetylcholinesterase. Finally, al neuron-to-neuron synapses, a single action potential on a presynaptic neuron rarely results in a full-blown action potential on the postsynaptic neuron. Some form of summation Ofpresynaplic inputs is required to generate a postsynaptic action potential.
As discussed in Chapter 6, action potentials on the muscle cell membrane lead to contraction, or mechanical shortening, of the muscle cell. When this contraction is combined with the shortening of many muscle cells, movement of the body occurs.