Therumen
Rumen acts as a huge fermentation vat that encompasses the entire left side of the animal’s abdominal cavity. The rumen, also known as the “paunch,” is lined with papillae for nutrient absorption and divided by muscular pillars into sacs.
The rumen is divided into dorsal and ventral sacs by an incomplete partition formed by the cranial and caudal pillars and the right and left longitudinal pillars. The dorsal part is divided into the cranial sac lying between the ruminoreticular fold and the cranial pillar, the dorsal sac, and the caudodorsal blind sac. Approximately 50 to 65 percent of starch and soluble sugar consumed is digested in the rumen. Rumen microorganisms (including bacteria, protozoa, and fungi) digest cellulose from plant cell walls, digest complex starch, synthesize protein from non-protein nitrogen, and synthesize B vitamins and vitamin K. These microorganisms use chemical and physical means of breaking down feed particles into simpler nutrients, thereby producing nutrient forms that both they and the animal need. Rumen pH typically ranges from 6.5 to 6.8. The rumen environment is anaerobic. One major product of carbohydrate fermentation is volatile fatty acids, which provide the animal with most of its energy. The wall of the rumen isDOI: 10.1201/9781003426851-16
147
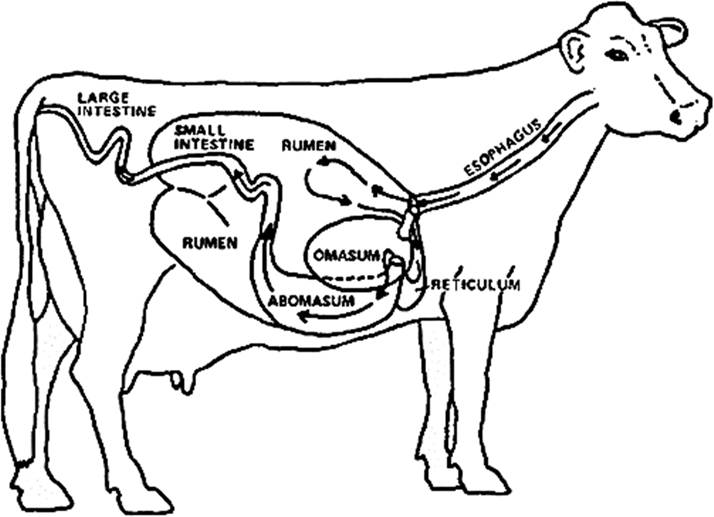
FIGURE 16.1 The figure of the ruminant digestive system describes different stomach parts like the rumen, reticulum, and omasum, as well as small and large intestine.
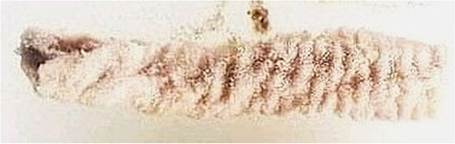
FIGURE 16.2 Rumen/Paunch is lined with papillae for nutrient absorption and divided by muscular pillars into sacs and acts as a fermentation vat.
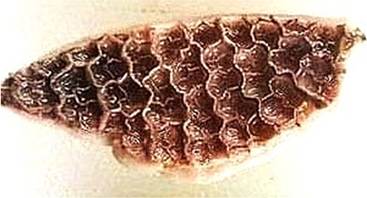
FIGURE 16.3 Figure of reticulum depicting the “honeycomb” network of low hexagonal ridges.
lined with small, fingerlike projections that are the primary site of absorption for volatile fatty acids. Rumen microorganisms also break down the portion of protein that is susceptible to fermentation and use it as building blocks to synthesize their own amino acids, which are incorporated into the microorganisms. These microorganisms are also capable of synthesizing amino acids from sources of nitrogen that are not true protein, commonly referred to as nonprotein nitrogen (Figure 16.2).
In addition to yielding nutrients that are beneficial to the animal, fermentation produces gaseous by-products, such as carbon dioxide, methane, ammonia, and hydrogen sulfide, that must be expelled by the animal. These gases are expelled through a process known as eructation that give the reticulum its common name, the “honeycomb” network of low hexagonal ridges. These ridges are papil- lated in sheep but smooth in cattle (Figure 16.3).
Foreign objects such as wire or nails that are swallowed typically will fall into and remain in the reticulum; contractions of this part of the forestomach may drive sharp objects through the wall of the stomach, leading to traumatic peritonitis or hardware disease. The reticular groove runs ventrally from the cardia to the reticulo-omasal orifice. In young ruminants, the lips of this groove can be brought into contact by contraction of the underlying muscle, so that a tube is formed by which the ingested milk bypasses the ruminoreticulum and directly enters the abomasum for the digestion.
16.3.1 The Reticulum
16.3.2 The Omasum
The reticulum is the most cranial compartment of the fore stomach. Its mucosa is thrown up into intersecting ridges Omasum is dense, kidney-shaped organ whose main function is the absorption of water and water-soluble nutrients.
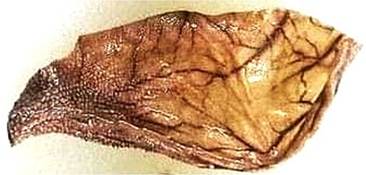
FIGURE 16.4 Omasum is a dense, kidney-shaped organ known as “many plies” due to the presence of many folds or leaves for increasing the surface area, which increases the area that absorbs nutrients from the feed and water.
It is relatively larger in larger ruminants like cattle and buffalo compared to smaller ruminants. It is called the “many plies” due to the presence of many folds or leaves for increasing the surface area, which increases the area that absorbs nutrients from the feed and water. Water absorption occurs in the omasum. It is connected to the reticulum by a short tunnel. After regurgitation of a bolus, it is swallowed and further subjected to ruminal fermentation (Figure 16.4).
16.3.3 The Abomasum
Abomasum also known as the “true stomach,” is quite similar to a stomach in a non-ruminant. The abomasum produces hydrochloric acid and digestive enzymes such as pepsin (which breaks down proteins) and receives digestive enzymes secreted from the pancreas, such as pancreatic lipase (which breaks down fats). The digesta undergoes acid and enzymatic digestion and then moves on to the small intestine. These secretions help prepare proteins for absorption in the intestines. The pH in the abomasum generally ranges from 3.5 to 4.0. The abomasum secretes mucus to protect the its wall from acid damage (Figure 16.5).
FIGURE 16.5 Abomasum or the “true stomach” is quite similar to a stomach in a non-ruminant dur to presence of digestive glands.
16.3.4 Pregastric Physiology
The organs of prehension vary somewhat in ruminants. In some, as in the cow, the lips, though strong, are not sufficiently movable to be of any use in prehension, while the tongue is so long and the muscles by which it is moved are so powerful that it is the principal organ used in this process. Sheep are grazers; they ramble slowly, eating short plants close to the ground. Goats are browsers and have clefted upper lips; they look for leaves, twigs, vines, and shrubs. A ruminant uses its mouth and tongue to harvest forages during grazing or to consume harvested feedstuffs. Cattle harvest forages during grazing by wrapping their tongue around the plants and then pulling to tear the forage for consumption.
They typically spend over one-third of their time grazing, one-third of their time ruminating (cud chewing), and slightly less than one-third of their time idling, where they are neither grazing nor ruminating.The roof of the ruminant mouth is a hard/soft palate without incisors. The lower jaw incisors work against this hard dental pad. The incisors of grass/roughage selectors are wide with a shovel-shaped crown, whereas those of concentrate selectors are narrower and chisel-shaped. Premolars and molars match between upper and lower jaws. These teeth crush and grind plant material during initial chewing and rumination.
Saliva is the mixed secretion of three pairs of main salivary glands, namely the parotid, submaxillary or submandibular, and sublingual of the mouth. These three glands are the major salivary glands in ruminants. In addition, sheep and cattle have two inferior molar glands and small, numerous buccal and labial glands in the cheeks and lips. Saliva aids in chewing and swallowing, contains enzymes for the breakdown of fat and starch, and is involved in nitrogen recycling to the rumen. When food enters the mouth, a copious secretion of saliva takes place by reflex stimulation of the salivary glands through the buccal receptors and secretory centers. Normal foodstuffs cause salivary secretion rich in mucin and enzymes to facilitate easy swallowing, while dry or noxious materials cause watery secretion with little mucin. The types and volumes of saliva secretion of saliva vary with a diverse range of stimuli. Saliva has a lubricating function that facilitates the mastication and swallowing of ingested food. The mucin content provides adhesion to food to form a bolus for swallowing. Spontaneous secretion of saliva, even during sleep, has an important protective function by keeping the oral mucosa moist and lubricated even in the absence of food. Salivary amylase is absent in horses, cattle, sheep, and goats. Saliva's most important function is to buffer pH levels in the reticulum and rumen.
In ruminants, saliva provides a proper medium for bacterial growth and activity in the rumen. The HCO3 and PO4 content of saliva neutralizes the volatile fatty acids produced during microbial fermentation. Urea, a non-protein source, supplies nitrogen for bacterial growth and microbial protein synthesis. A mature cow produces up to 160-200 liters of saliva per day, but this varies depending on the amount of time spent chewing feed, as that stimulates saliva production. Forage and feed mix with saliva containing sodium, potassium, phosphate, bicarbonate, and urea when consumed to form a bolus.That bolus then moves from the mouth to the reticulum through a tube-like passage called the esophagus. Muscle contractions and pressure differences carry these substances down the esophagus to the reticulum. Ruminants eat rapidly, swallowing much of their feedstuffs without chewing them sufficiently (< 1.5 inches). The esophagus functions bi-directionally in ruminants, allowing them to regurgitate their cud for further chewing, if necessary.
The process of rumination or “chewing the cud” is when forage and other feedstuffs are forced back to the mouth for further chewing and mixing with saliva. This cud is then reswallowed and passed into the reticulum. Then the solid portion slowly moves into the rumen for fermentation while most of the liquid portion rapidly moves from the reticu- lorumen into the omasum and then the abomasum. The solid portion left behind in the rumen typically remains for up to 48 hours and forms a dense mat in the rumen where microbes can use the fibrous feedstuffs to make precursors for energy.
16.3.5 Fermentative Digestion in Rumen
Ruminal fermentation initially results in the degradation of carbohydrates and proteins to short-term intermediates such as sugars and amino acids. The products of this initial degradation are readily metabolized to microbial mass and carbon dioxide, methane, ammonia, and volatile fatty acids (VFA): primarily acetate, propionate, and butyrate, and to a lesser degree branched chain VFA and occasionally lactate.
The rate and extent of fermentation are important parameters that determine protein, vitamins, and short-chain organic acids supply to the animal (Koenig et al., 2003). The host ruminant animal absorbs VFA (mostly through the rumen wall) and digests proteins, lipids, and carbohydrate constituents of microbes and feed residues entering the small intestine to supply its maintenance needs and for the production of meat and milk. Ruminant animals derive about 70% of their metabolic energy from microbial fermentation of feed particles and microbial protein accounts for as much as 90% of the amino acids.Rumen microorganisms usually adhere to feed particles and form biofilms to degrades plant material. The efficiency of ruminants to utilize of feeds is due to highly diversified rumen microbial ecosystem consisting of bacteria (1010-1011 cells/ml, more than 50 genera), ciliate protozoa (104-106 / ml, 25 genera), anaerobic fungi (103- 105 zoospores/ml, 5 genera) and bacteriophages (108-109 /ml). The synergism and antagonism among the different groups of microbes is so diverse and complicated that it is difficult to quantify the role played by any particular group of microbes present in the rumen (Kamra, 2005). Bacterial numbers in the rumen are the highest and bacteria play a dominant role in all facets of ruminal fermentation.
16.3.6 Some Advantages of the Fermentation
in the Reticulorumen
1. Allows digestion and then absorption of fermentation products that are of value to the host (e.g., microbial cells, VFAs, and B vitamins) before the acidic abomasum.
2. May change poor quality protein and other dietary nitrogen compounds to good-quality microbial protein.
3. Selective retention of coarse particles extends fermentation time and allows for further mechanical breakdown during rumination (cud chewing).
4. Release of fermentation gas (mostly CO2 & CH4) from the system by eructation.
5. Toxic substances in the diet may be attacked by the microbes before being presented to the small intestine.
16.3.7 Eructation
It is the expulsion of fermentation gases like CO2 and methane that has accumulated in the rumen. Receptors for eructation reflex are the tension receptors located in the reticulum, cardia, and cranial rumen sac, which are stimulated by accumulation of gas. Centre for eructation reflex are present in Medulla Oblongata. Certain amount of gas is produced in ruminant animal due to fermentation process. The gas layer is moved cranially by the secondary contraction of the dorsal sac. From there it enters into the cardia and then lower oesophageal sphincter will open, and the gas enters into the relaxed oesophagus. By an antiperistal- tic wave in the oesophagus and elevation of soft palate the gas is expelled through the mouth. Eructation occurs once in two ruminoreticular contractions. Inhibition of eructation is observed during blokage of ruminoreticular contraction. Blockage of eructation leads to bloating of the rumen, called as tympany which can also takes place when gas production is higher than its elimination. Distension of rumen may cause hindrance of respiratory process as diaphragm did not expand and contract easily. Oils and non absorbable surfactants are used to treat bloat. High grain feeding and feeding of lush alfalfa results in trapping of gases in the rumen called feedlot bloat. This is due to formation of foam in the ruminoreticulum which traps many small gas bubbles. This can be prevented by reducing feeding of grains or lush green pastures.
16.3.8 Ruminal-Reticular Contractions
There are mainly two patterns of motility seen in reticulo- rumen as primary or mixing contractions called primary cycle and secondary or eructation contractions known as secondary cycle. In the primary contraction, rumino- reticular contractions begin from the reticulum as biphasic contractions of which the first contraction known as mixing contraction. It relaxes for sometime and then again the second contraction with strong force is called as evacuation contraction begin. The function of the secondary contraction is to expel the gases out of the rumen. The reticulo- rumen contractions occur at a frequency of 1 to 3/ min. It may increase during feeding. Third contraction follows the second strong contraction during rumination. This third contraction is the strongest of the three and is referred to as extra or regurgitation contraction of the reticulum.
16.3.9 Microbial Digestion (Role of
Bacteria, Fungi, and Protozoa)
Dietary carbohydrates are the main rumen microbial fermentation substrates. Microbial yields are related primarily to the growth rate that carbohydrate permits. The individual carbohydrates characterized by faster rumen degradation rates result in greater microbial yield (Hall and Herejk, 2001). The enzyme systems produced by microorganisms for carbohydrates hydrolysis are complex; they usually comprise hydrolases from several families, and there may be multiple enzymes hydrolysing each polysaccharide. Nearly all carbohydrate digestion occurs (>90%) within the rumen, but under certain circumstances (e.g., high rate of passage), a significant amount of carbohydrate digestion can occur in the small and large intestine. Carbohydrate digestion in ruminant animals is through microbial fermentation in the rumen. Dietary carbohydrates are degraded (fermented) by rumen microbes (bacteria, fungi, protozoa). The purpose of rumen fermentation is to produce energy as ATP for the bacteria to use for protein synthesis and their own growth. VFAs, also known as short-chain fatty acids, are produced as a product of rumen fermentation and are absorbed through the rumen wall and are utilized by the animal as an energy source. In young ruminants, rumen and the reticulum are not fully developed and are relatively small. The reticular/oesophageal groove reflex, a tube-like fold of tissue, channels milk or water that is sucked from a nipple directly through the omasum to the abomasum. This is a reflex, stimulated by sucking. When the animal is weaned, it normally loses this reflex. Solid food, such as creep feed, passes into the small rumen and fermentation starts. The neonatal ruminant animal has no ruminal bacterial population but from birth, it starts to pick up bacteria from the mother and environment, particularly through contact. Solid food is then fermented forming VFAs, which stimulate the growth and development of the rumen, particularly the growth of the papillae for absorption.
Cellulolytic bacteria tend to produce more acetate, while amylolytic bacteria produce more propionic acid. Typically, three major VFA molar ratios are 65:25:10 with a roughage diet and 50:40:10 with a concentrate-rich diet. Changes in VFA concentration can lead to several disorders of carbohydrate digestion in ruminants. Rumen acidosis occurs when animals are fed high-grain-rich diets or when animals are suddenly changed from pasture- or range-fed to feedlot conditions. Cellulose content is in the range from 35 to 50% of plant dry weight. It is chemically composed of a homogenous polymer of β-1,4-D-glucose linked through β-1,4- glycosidic bonds. Fibrobacter succinogenes, Ruminococcus flavefaciens and R. albus are considered to be the predominant cellulolytic bacteria present in the rumens, these species gain selective advantage in the rumen is by optimizing two catabolic activities: cellulose hydrolysis (depolymerization) and efficient utilization of the hydrolytic products (cellodextrins) (Koike and Kobayashi, 2001). F. succinogenes has a potent ability to solubilize crystalline chains of cellulose (Shinkai and Kobayashi, 2007). F. succi- nogenes produces primarily succinate (a propionate precursor), and lesser amounts of acetate, R. flavefaciens produces primarily acetate and lesser amounts of succinate converts to propionate by Selenomonas ruminantium (Weimer et al., 1999).
The amylolytic bacteria such as Streptococcus and Ruminobacter are particularly adapted for digesting starch and sugar. Under the strict anaerobic conditions within the rumen these bacteria ferment the starches and sugars to lactic acid, along with some VFAs. The ruminants which are generally high grain fed, the action of the amylolytic bacteria can be a particular concern to those ruminants. The amylolytic bacterial populations can multiply very rapidly in response to the starch in the diet and these bacteria produces large amounts of lactic acid. Lactic acid is nearly 10 times stronger acid than acetate, propionate and butyrate. Due to high amounts of lactic acid and other VFAs in the rumen fluid, pH of rumen fluid fall below 5.7. Because of this pH the cellulolytic bacteria begin to die and also the rumen epithelium can be damaged by the accumulation of large amount of acid. As the bacteria dies, the dying bacteria release endotoxins that enter the blood and can cause shock. This condition is known as rumen acidosis. This type of condition mostly occur in cattle that have not been allowed time to acclimatize to the high grain diet. If the amount of grain in the diet is increased slowly over a period of weeks, this type of condition like ruminal acidosis does not develop because it allows populations of bacteria known as lactate utilizers to populate the rumen. These bacteria metabolize the rumen fluid lactate as an energy source. The lactate utilizers bacteria belong to the Selenomonas and Megasphaera genera. Animals with very high populations of these bacteria in the rumen can be fed very high starch diets with little risk of rumen acidosis because acclimatization already been takes place. Hindgut fermenters like horses, rabbits and others have essentially the same types of cellulolytic bacteria living in their cecum and colon as cows have in their rumen. However, in these species starches and sugars are absorbed by the small intestine before they reach the colon so the risk for very low pH is reduced.
In addition to a wide variety bacteria, the rumen also contains small populations of fungi. Some of these fungal species may help degrade lignin, a woody and indigestible constituent of plant cell walls. Other inhabitants of the rumen are the protozoa. The protozoa feed on ruminal bacteria, plant starch granules and other readily digestible materials including polyunsaturated fatty acids. Most of the protozoa are located in the raft of fibrous digesta in the dorsal ruminal sac and play many important functions like it form a reservoir of microbial protein useful at time intermittent food supply, also helps in prevention of overpopulation of bacteria, also made available polyunsaturated fatty acids to the lower part of GI tract for their absorption. Protozoa are very sensitive to change in pH of the rumen. The protozoa and primary cellulolytic bacteria require pH
6.2 or greater, whereas the primary amylolytic bacteria require slightly acidic pH of around 5.8.
16.3.10 Microbial Protein
The rumen bacteria used to provide very high quality protein to the animal because its amino acid profile is almost identical to that of muscle and milk, permitting great conversion into meat and milk by the cow. Ruminal bacteria have the ability to form all the essential amino acids from the available resource. When these bacteria die, the proteins within the bacteria can be digested by mammalian proteolytic enzymes and the available amino acids are used by the ruminant animal. Two types of protein are found in ruminant animal, rumen-degradable protein and rumen-nonde- gradable protein. Dietary protein that the rumen bacteria can break down is referred to as rumen-degradable protein. It is essential to feed high quality proteins to manogastric animals to supply essential amino acids because these animals cannot synthesis high quality protein. In ruminants, if the protein is rumen degradable the essential amino acids will be lost and it will be of no use unless they can be recovered in the form of microbial protein that enters the small intestine. All the dietary protein fed to a cow is not degraded by the rumen bacteria for their use. The protein that bypasses the rumen bacteria, known as rumen-nonde- gradable protein. These protein can be digested in the small intestine and can be an excellent source of essential amino acids, if it is of high quality.
16.3.11 Formation of Volatile Fatty
Acids and Their Absorption
The most important end products of carbohydrates fermentation in ruminants is volatile fatty acids. These volatile fatty acids are the major source of energy for the ruminants. These include acetic acid, butyric acid and propionic acid. Fat and protein contents of milk are also determine by these volatile fatty acids. After their production, these volatile fatty acids are absorbed through the walls of rumen and transported via blood to the liver. The granulosa cells of the forestomach epithelium contain carbonic anhydrase, which promotes the formation of carbonic acids (Figure 16.6).
This carbonic acids are dissociates into bicarbonate ions and hydrogen ions. The hydrogen ions associate with volatile fatty acid anions to form undissociated VFAs that can diffuse more easily across the epithelium, leaving bicarbonate ions in the ruminal fluid. This mechanism also reduces
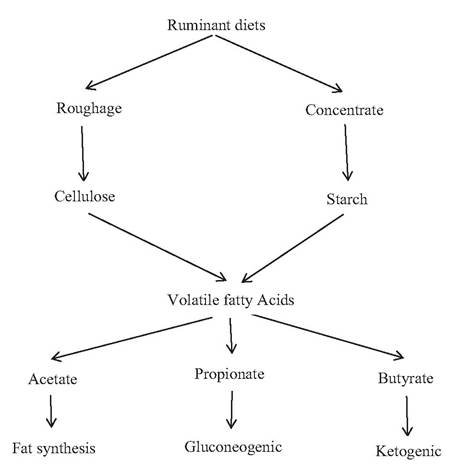
FIGURE 16.6 Formation of volatile fatty acids and their absorption.
ruminal acidity by exchanging the anions of stronger acids for those of a weaker acid (carbonic acid). Most of the acetate in liver is converted to acetyl CoA via acetyl CoA synthetase. Most of the acetate (approximately 80%) escapes oxidation in liver and is oxidized by citric acid cycle or used for fatty acid acids synthesis. These acetate is the main precursor for lipogenesis in ruminants. Higher activity of acetyl CoA synthetase in ruminant adipose tissue reflects the high rate of acetate conversion to fatty acid. Acetate is also the main precursor for milk fat synthesis. Approximately around 2-5% of propionic acid is converted to lactic acid during its absorption through the rumen,while the remainder enters in the portal blood as propionate. Most of the propionic acid is oxidized to glucose in liver. Propionyl CoA undergoes CO2 fixation by the biotin containing enzyme, propionyl CoA carboxylase, to form methylmelonyl CoA and subsequently converted to succinyl CoA. This succinyl CoA then enters the citric acid cycle for the production of ATP as source of energy for the ruminants. Butyrate during its absorption through the ruminal epithelium is converted to ketones and is present in low level in portal blood. More than 80% of the ketones formed occurs in the form of β-hydroxybutyric, whereas the remaining comprised of acetoacetate and acetone. The β-hydroxybutyric acid is mainly used for fatty acid synthesis in adipose and mammary gland tissue, whereas it is oxidized in cardiac and skeletal muscles. Butyrate that is reaching the liver is rapidly metabolized by hepatic tissue.
16.3.12 Importance of FVAs in Ruminants
VFAs play very important and significant role in ruminant animal. Acetate is the precursor for milk fat synthesis. Propionate is glucogenic acids and provides glucose through gluconeogenesis. Acetate and butyrate contribute the energy needs of the ruminant animal. These fatty acids provide energy by entering Krebs’s cycle as different substrate. Butyrate and acetates are mainly ketogenic acids and causes the formation of the ketone bodies like acetone, acetoacetic acid and beta hydroxy butyric acid. The ketone bodies are an important source of energy in tissues like brain and heart.
16.3.13 Other End Products of
Fermentation in Ruminants
Lactic acid is a stronger acid than the volatile fatty acids. Certain amylolytic bacteria produce lactic acid along with volatile fatty acid during degradation of starch. Generally lactic acid found in both isomeric form and occurs as a transient fermentation intermediate. With high starch diets, it may accumulate in the forestomach with low pH conditions, initially only slowly absorbed but later largely converted to glucose in the liver. The principal gases produced by ruminant animal after fermentation of substrate is carbon dioxide (60%) methane (30-40%) and variable amount of nitrogen, with trace amount of H2S, hydrogen and oxygen. Carbon dioxide is produced by decarboxylation reactions of fermentation and from the neutralization of volatile fatty acid hydrogen ions by bicarbonate entering the rumen in saliva. Methane is produced from the reduction of carbon dioxide and formate by methanogenic bacteria. Hydrogen sulfide is produced by the reduction of sulfate and sulfur containing amino acids. This gas is generally toxic even in small quantity. Ruminal environment is generally low in oxygen concentration and sometimes oxygen may enter the rumen through trapped with ingested food and water and also sometimes by diffusion from blood but it is quickly used by the facultative anaerobic bacteria. Ammonia is produced by the deamination of dietary proteins, from non protein nitrogenous substance (NPN) and from urea derived from saliva. Urea feeding is beneficial to the animal and it is always advisable to feed urea as nitrogen supplement (feeding upto 30% total nitrogen as a urea supplement).
16.3.14 Nutrient Reabsorption in Small Intestine
The small intestine is the main site of absorption of nutrients that have by-passed the rumen. Bile and pancreatic juice drain into the small intestine to aid in digestion of certain feed components, such as dietary fat. The liver is a large organ, about 1.5% of an animal’s live weight. It is located towards the front of the animal’s body cavity, just behind the diaphragm. Newly digested and absorbed nutrients are transported from the absorption sites to the liver for storage and/or further processing. The liver also produces bile that drains into the intestine to aid in the digestion of fats.