Urinary System
Avian kidneys lie on the ventral surface of the synsacrum. Each kidney is divided into cranial, middle, and caudal lobes created by external grooves associated with large vessels that cross the renal surface (Fig.
30-6). Collecting ducts empty directly into the ureter adjacent to the cranial lobe of the kidney; there is neither renal pelvis nor renal calyces as in other domestic species. Right and left ureters terminate in the dorsal part of the urodeum.uniquely, birds possess a renal portal system in addition to the expected renal arterial blood supply (one renal artery per lobe) and renal venous return (Fig. 30-6). This system, consisting of cranial and caudal renal portal veins, receives venous blood from the caudal body and distributes it into a second capillary system in the interlobular spaces.
The presence of the renal portal system has been used as a justification for avoiding the caudal body as a site for injection of medications in birds. One reason for this avoidance is that the drugs may be more rapidly excreted when injected here than is desirable due to their being immediately transported to and removed by the kidney. Another concern is that drugs that are potentially nephrotoxic (damaging to the kidneys) may be more so when they are first distributed undiluted to the kidneys via the portal system.
As in mammals, avian kidneys are composed of nephrons that perform glomerular filtration, selective tubular reabsorption, and selective tubular secretion in the process of urine formation. However, avian kidneys contain two different types of nephrons, and one type is markedly different in structure compared to nephrons of mammalian kidneys. This unique type of nephron is termed a reptilian-type (RT) nephron and is similar to those found in kidneys of reptiles. Renal corpuscles of RT nephrons are found in the lobules of avian kidneys, and a renal tubule connects each renal corpuscle to a collecting duct also found in the lobule (Fig.
30-7). However, the tubules of the RT nephrons do not include a segment that corresponds to a loop of Henle, and the tubules of the RT nephrons are contained in the outer zones of the lobules, where they connect to collecting ducts (Figure 30-7). Avian kidneys also contain mammalian-type (MT) nephrons that do have segments that correspond to the loops of Henle in mammalian kidneys, and these loops extend downward into medullary cones that support the lobules (Figure 30-7). These tubular loops of MT nephrons in avian kidneys function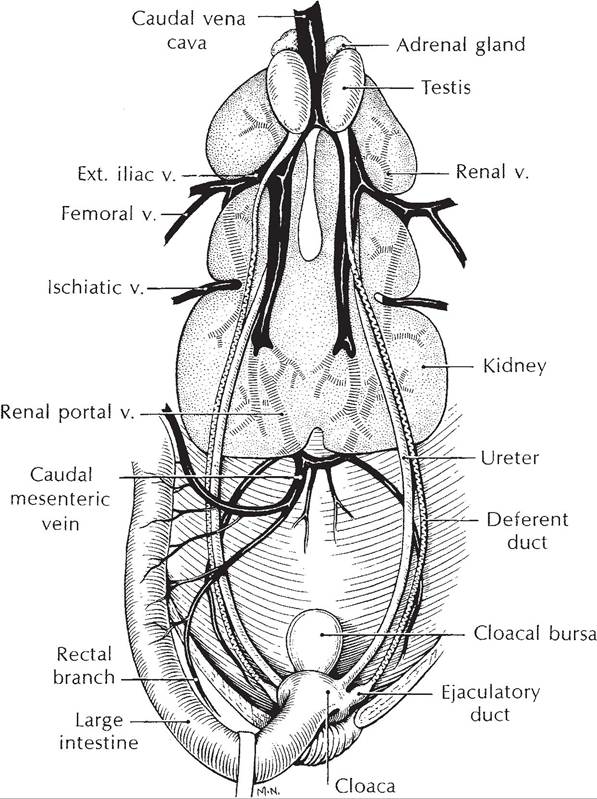
Figure 30-6. The avian genitourinary tract (male).(Reprinted with permission of Wolters Kluwer from Rosskopf,
W. and Woerpel, R. Diseases of Cage and Aviary Birds, 3rd ed. Philadelphia: Williams & Wilkins, 1996.)
similarly to loops of Henle in mammalian kidneys and contribute to the formation of an osmotic gradient within the interstitial fluids of the medullary cone. However, the magnitude of the gradient is much less in birds. Tubular segments corresponding to proximal tubules and distal tubules can be found in both MT and RT nephrons, and these segments in birds perform secretory and reabsorptive functions similar to nephrons in mammalian kidneys.
Glomerular filtration rate (GFR) remains relatively stable in normal mammals despite mild to moderate changes in hydration status. However, in birds, moderate dehydration, such as with periods of water restriction, is associated with significant reductions in GFR. Some suggest that this is primarily due to vasoconstriction of vessels supplying the RT nephrons.
To affect water balance in response to changes in water intake, the water permeability of collecting ducts in mammalian kidneys is regulated by antidiuretic hormone. Collecting ducts in avian kidneys respond similarly to arginine vasotocin (AVT), the avian antidiuretic
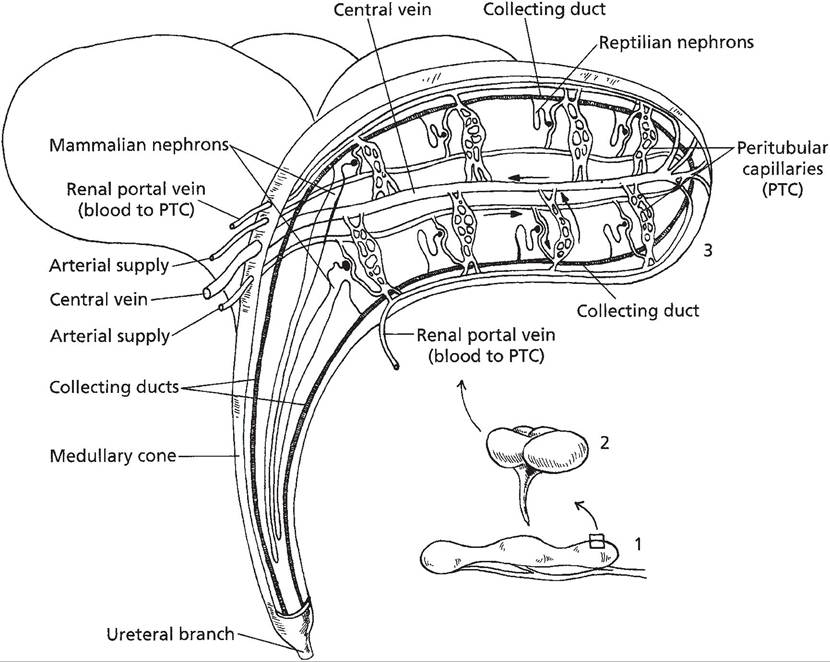
Figure 30-7.
Location of mammalian and reptilian type nephrons within a lobule of a typical avian kidney. Renal portal veins serve as a secondary blood supply to peritubular capillaries. (Reprinted with permission of Wiley- Blackwell from Reece, W.O. Functional Anatomy and Physiology of Domestic Animals, 3rd edition. Baltimore: Lippincott, Williams & Wilkins, 2005.)hormone, and increase their water permeability. The release of AVT from the posterior pituitary responds to changes in extracellular fluid osmolality in a manner similar to that for mammals. When water intake is excessive, AVT levels are reduced and a more dilute urine is excreted; when water intake is restricted, AVT levels increase and a more concentrated urine is excreted. However, because the maximal osmolality of the interstitial fluid surrounding collecting ducts is less in birds than in mammals, maximal urine osmolality is also less.
Birds also have other means to promote water reabsorption from renal tubules, and some of these means are related to the excretion of nitrogenous wastes. Both mammals and birds eliminate nitrogenous wastes through urine, but the chemical form of the waste differs between mammals and birds. Mammals primarily excrete urea, whereas birds primarily excrete uric acid. Urea enters tubular fluid (potential urine) of mammals by glomerular filtration. Uric acid enters tubule fluid in avian kidneys by both glomerular filtration and tubular secretion with secretion greater in most cases. The unique renal portal system described above supplies blood to a peritubular capillary network around tubules and is a source of uric acid for secretion, even when glomerular blood flow and glomerular filtration are reduced.
Because urea is not a major nitrogenous waste in birds, measure of blood urea nitrogen (BUN) levels cannot be used as indicators of renal function. Plasma uric acid levels can be used, but because most uric acid enters tubular fluid by secretion, significant renal tubular dysfunction must occur before plasma uric acid increases.
Within renal tubules of avian kidneys, unique, extremely small (3-13 μm), spherical concretions or particles form, and these contain uric acid, albumin, and inorganic ions. The accumulation of multiple osmotic particles into a single particle effectively reduces the osmolality of the tubular fluid and permits additional water reabsorption. Albumin is readily available in the tubular fluid of birds for it contains significantly more plasma protein than in mammals.
As discussed earlier, urine may be moved from the urodeum by reverse peristalsis to the ceca, and additional absorption of water and electrolytes may occur in the coprodeum, rectum, and ceca. Uric acid in the urine entering the cloaca may precipitate on feces, and these are seen as a white pasty part of the droppings.