Ventilation Requires Muscular Energy
Energy provided by muscles causes air to enter the lungs during inhalation. During exhalation, much of the energy causing air to leave the lungs is provided by the elastic energy stored in the stretched lung and thorax.
Therefore, in most animals at rest, inhalation is an active process, whereas exhalation is passive. Horses are an exception to this general rule because they have an active phase to exhalation even at rest. During exercise or in the presence of respiratory disease, exhalation often is assisted by muscle contraction in most species.The primary inspiratory muscle is the diaphragm, which is a domed musculotendinous sheet separating the abdomen and thorax and innervated by the phrenic nerve. The diaphragm consists of a costal portion, arising from the xiphoid process and the costochondral junctions of the 8th to 12th ribs (8th to 14th ribs in Equidae), and a crural portion, arising from the ventral surface of the first three to four lumbar vertebrae and extending toward the tendinous center of the diaphragm. The apex of the dome of the diaphragm extends TOStrally to the seventh or eighth intercostal space at the level of the base of the heart. During contraction of the diaphragm the dome is pulled caudally and thereby enlarges the thoracic cavity. The tendinous center pushes against the abdominal contents, elevating intraabdominal pressure, which displaces the abdominal wall and caudal ribs outward, thus also tending
to enlarge the thorax. It is the enlargement of the thorax that creates the negative (subatmospheric) pressure necessary to make air flow into the lungs during inhalation.
The external intercostal muscles also are active during inhalation. The fibers of these muscles are directed Caudoventrally, from the caudal border of one rib to the cranial border of the next, so that muscle contraction moves the ribs rostrally and outward.
The relative contributions of diaphragmatic and costal movement to ventilation under different metabolic demands are not well defined in animals. Because the cranial ribs support the forelimbs in quadrupeds, they participate less in ventilation than do the more caudal ribs. Other inspiratory muscles include those connecting the sternum and head. These muscles contract during strenuous breathing and move the sternum rostrally.The subatmospheric pressure generated within the respiratory tract during inhalation tends to collapse the external nares, pharynx, and larynx. Contraction of abductor muscles attached to these structures is essential for preventing collapse. Abductor muscle contraction during inhalation can be observed as dilation of the external nares. Laryngeal hemiplegia in horses is a condition in which the abductor muscles on the left side of the larynx fail to contract during inhalation. During exercise these horses exhibit a sound known as “roaring.” Roaring occurs as the result of the turbulent airflow that is generated on inhalation as the vocal fold on the paralyzed side is sucked into the lumen of the larynx by the subatmospheric pressure.
The principal expiratory muscles are the abdominal muscles and internal intercostal muscles. Contraction of the abdominal muscles increases abdominal pressure, which forces the relaxed diaphragm forward and reduces the size of the thorax. The fibers of the internal intercostal muscles are directed cranio- ventrally, from the cranial border of one rib to the caudal border of the next cranial rib, so that their contraction decreases
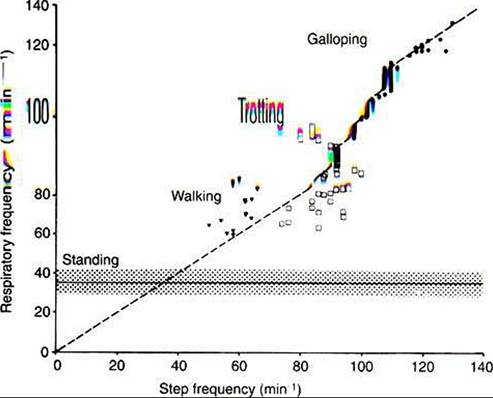
FIGURE 45-4 Relationship between gait and respiration in the horse. In the walk and trot, step and respiratory frequency are not correlated. At the gallop (and canter), respiratory and step frequency bear a 1:1 relationship. (Modified from Hornicke H, Meixner Rz Pollman U: EQuine exercise physiology, Cambridge, UK, 1983, Granta Editions.)
the size of the thorax by moving the ribs caudally and ventrally.
As the thorax becomes smaller, the thoracic pressure increases and forces air out of the lungs.During exercise, respiratory muscle activity increases in order to generate the increase in Ve. In cursorial (running) mammals, ventilation is synchronized with gait in the canter and gallop, but not in the trot (Figure 45-4). Inhalation occurs as the forelimbs are extended and the hind limbs are accelerating the animal forward. Exhalation occurs when the forelimbs are in contact with the ground. In the galloping horse and perhaps in other galloping quadrupeds, much of the increase in size of the thorax during inhalation is a consequence ofelongation of the trunk rather than an increase in the diameter of the thorax.
The Respiratory Muscles GenerateWork to Stretch the Lung and Overcome the Frictional Resistance to Airflow
At the end of a normal exhalation, some air (~45 mL∕kg) remains in the lung. This air volume is known as functional residual capacity (FRC). At FRC, the pressure in the pleural cavity (Ppl) that surrounds the lung is approximately 5 cm H2O below atmospheric pressure (-5 cm H2O). As previously mentioned, Ppl decreases during inhalation as the thorax enlarges and the respiratory muscles perform work to stretch the elastic lung and thorax and to generate airflow against the frictional resistance of the air passages (Figure 45-5). Lung compliance is a measure of the elastic properties of the lungs, and airway resistance is a measure of the frictional resistance of the airways. The magnitude of the change in pleural pressure (ΔPpl) during each breath is determined by the change in lung volume (ΔV), by lung compliance (C), by airflow rate (V), and by airway resistance (R), as follows:

Resting animals breathe slowly and have low flow rates. In this situation the primary work of the respiratory muscles is against the compliance of the lung.
When the respiratory rate
Lung Elasticity Results from Tissue and Surface Tension Forces
At FRC, the slightly Subatmospheric pressure in the pleural cavity keeps the lung inflated. If the thorax is opened and the lungs are exposed to atmospheric pressure, the lungs collapse to their minimal volume. At this volume, some air remains trapped within the alveoli behind closed bronchioles. This trapped gas causes collapsed normal lungs to float in water. The collapse of the lung when the thorax is opened is a result of the lungs inherent elasticity, which is generated by both elastic and collagen tissue and by surface tension forces.
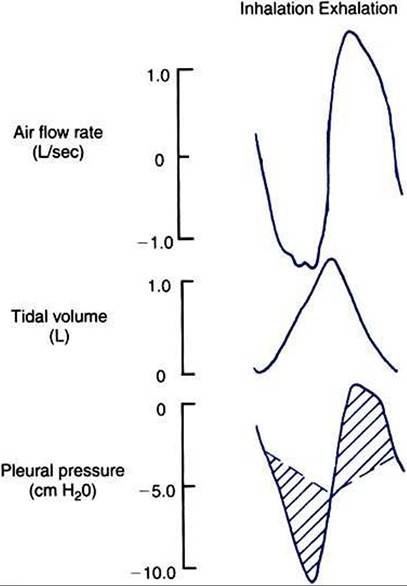
FIGURE 45-5 Airflow rate, tidal volume, and pleural pressure during inhalation (left) and exhalation (right). During inhalation, pleural pressure decreases as the thorax enlarges, air flow rates increase, and the volume of air in the lung increases. At the end of inhalation, flow rates return to zero, and pleural pressure becomes less negative. During exhalation, pleural pressure increases as the thorax decreases in size, flow rates increase to a peak and then decrease again, and the volume of air in the lung decreases. The broken line shows the change in pleural pressure necessary to overcome the frictional resistance of the airways.The peaks of pleural pressure on both inhalation and exhalation coincide with peaks of flow.
The importance of surface tension can be demonstrated experimentally by inflating the lung with air or saline while concurrently measuring the pressure required for inflation, also known as transpulmonary pressure (PL). Figure 45-6 represents the resulting lung pressu re-vol time curve and demonstrates the following important points:
1. A high pressure is required initially to inflate the lung with air from the gas-free state.
This is because high pressure is required for the initial opening of the bronchioles. In life, the lung is gas free only in the fetus and for a few seconds after birth until the first breath is taken. Usually, tidal breathing is initiated from FRC. At this lung volume, the bronchioles are open.2. The lung reaches its elastic limits at PL of approximately 30 cm H2O. The volume of air in the lung at this point is known as the total Iuny capacity.
3. The lung’s elastic properties differ during inflation and deflation; less pressure is necessary to maintain a given volume during deflation than during inflation. This phenomenon, known as pressure- volume hysteresis, is a result of changes in surface tension forces, the same forces that made it necessary for the high pressure initially to inflate the lung.
4. When saline is used instead of air to inflate the lung, less pressure is required for inflation, and the pressure-volume hysteresis is abolished. These two phenomena occur because when the lung is inflated with saline, the air-liquid interface of the fluid film overlying the alveolar epithelium is abolished. Because surface tension forces arise from this interface, they too are eliminated when the lung is inflated with saline. A comparison of the pressure-volume curves when the lung is inflated with air and with saline (see Figure 45-6) shows that surface forces are responsible for a considerable part of the elastic recoil of the air-filled lung. Surface tension forces continually try to collapse the
alveoli. Alveolar stability is a consequence of the presence of a pulmonary surfactant that reduces the surface tension of the alveolar lining. Pulmonary surfactant is a mixture of lipids
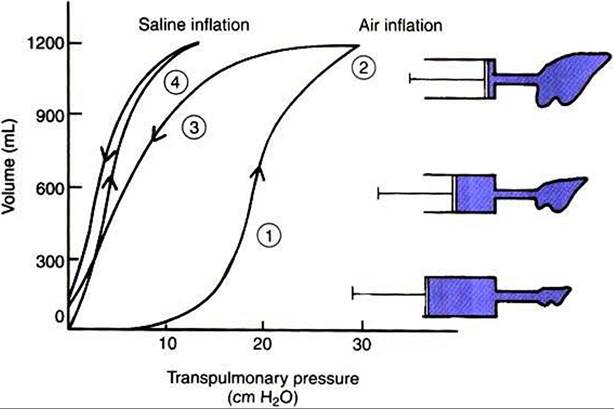
FIGURE 45-6 Pressure-volume curve of the lung during inflation with saline and with air. The pressure gradient across the lung (transpulmonary pressure) is shown on the abscissa and lung volume on the ordinate.
During air inflation the inflation and deflation curves are separated; that is, there is pressure-volume hysteresis. Hysteresis is abolished by inflating the lung with saline.The lung inflates more easily (i.e., it takes less pressure for a given volume) with saline than with air. Numbers correspond to numbers in the section Lung Elasticity Results from Tissue and SurfaceTension Forces.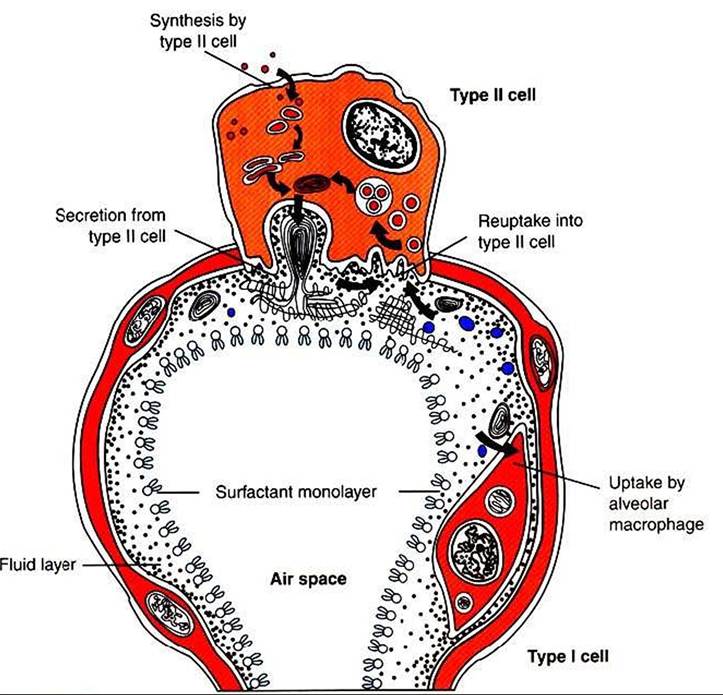
FIGURE 45-7 Diagram of an alveolus to show the movement of surfactant components through the type Il cell and the alveolar liquid.
and proteins. The most plentiful lipid component, dipalmitoyl phosphatidylcholine, is responsible for the surface tension reduction. Surfactant is produced in type II alveolar cells, and its hydrophilic and hydrophobic portions cause it to seek the surface of the alveolar lining (Figure 45-7). As lung volume decreases and the alveolar surface area shrinks, surfactant molecules become concentrated on the alveolar surface, reducing surface tension and promoting alveolar stability.
There are four important surfactant apoproteins. Two of these, surfactant proteins B and C, are hydrophobic and aid in surfactant secretion, maintenance of surface films, and reuptake of surfactant into type 11 cells. The other two apoproteins, surfactant proteins A and D, are hydrophilic and have antimicrobial properties.
Pulmonary surfactant is released into the alveolar spaces and tracheal fluid late in gestation (85% of the length of gestation in the sheep). Its appearance correlates with the rise in fetal plasma cortisol levels. Animals born prematurely have difficulty inflating their lungs because of inadequate surfactant. Synthetic surfactants can be used to treat premature newborns that lack adequate surfactant.
Lung compliance is the slope of the lung-pressure volume curve (see Figure 45-6). Because the pressure-volume curve is not linear, compliance obviously varies with the state of lung inflation. It is usually measured over the range of VT and, when adjusted for differences in lung size, does not vary greatly among adult mammals. Consequently, most mammals generate similar changes in pleural pressure during breathing. Anesthesiologists frequently refer to lung compliance when trying to artificially ventilate an animal. A “compliant lung” is easy to inflate. A “lung with low compliance,” as occurs in some diseases, is difficult to inflate.