VISION
1. Identify the parts of the external eye.
2. Identify the basic structures of the eyeball.
3. What is the normal arrangement of the corneal stroma?
4. Does the cornea have a blood supply?
5.
Are there nerve fibers in the cornea?6. How is accommodation accomplished by the lens of the eye?
7. What is the extent of accommodation among domestic animals?
8. How is the size of the pupil changed?
9. Are pupil shapes the same among all animals?
0. What would be the impact of a centrally located optic disk and a constricted circular pupil?
11. Review the production, location, circulation, function, and drainage of aqueous humor.
2. What visual chemical when excited by light begins to decompose and stimulates retinal rod cells?
3. What is the tapetum and how does it provide for better vision in reduced light?
4. What is meant by the field of vision, by binocular vision, and by monocular vision?
L5. Describe the conjunctiva.
6. If the nasolacrimal duct were plugged, explain the reason for a horse’s wet face.
L7. What is a “cherry eye” in the dog?
The receptor organs for vision are the eyes. The receptor stimulus is light and, accordingly, many of the eye structures are adapted for transparency so that light rays can reach the receptors. The parts of the eye as seen from the front (the external eye) are shown in Figure 5-16. The basic structures of the eyeball are shown in Figure 5-17.
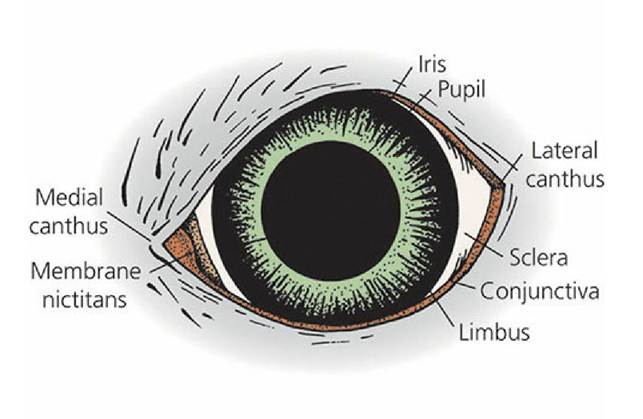
■ FIGURE 5-16 The external eye. The medial canthus is on the nasal side. The nasolacrimal duct originates in the medial canthus. The limbus is the junction of the sclera with the iris.
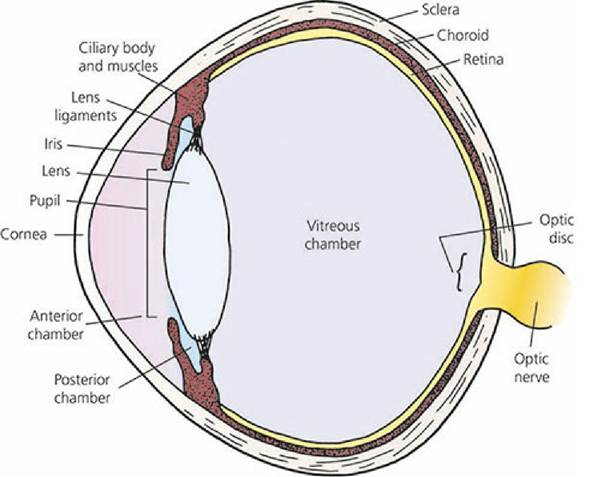
■ FIGURE 5-17 Diagram of an eyeball showing its basic structure.
The pupil is the opening between the central projection of the iris.Structure and Functions of the Eye
The eye is composed of the eyeball (globe), the optic nerve, and the accessory structures, which comprise the eyelids, conjunctivae, lacrimal apparatus, and the muscles of the eyeball.
Tunics of the Eyeball
The eyeball has three distinct layers or coats, known as tunics. The outermost external coat (fibrous tunic) is the supporting layer of the eyeball and is composed of the anterior cornea and posterior sclera. The sclera is the tough, white part of the fibrous tunic. The middle coat is the vascular tunic and is composed of the choroid, ciliary body, and iris. The innermost tunic is the light-sensitive retina (nerve tunic). It consists of several layers; three of its layers are cells. The light-sensitive cell layer consists of the rods (black-and-white vision) and the cones (color vision). These receptors convert light to a nerve impulse. The retina is black because of the presence of melanin. This black pigmentation not only assists in the absorption of light but also prevents uncontrolled reflection to other parts of the eye.
Cornea
The cornea is the transparent, forward continuation of the sclera. It is transparent to allow for the entrance of light. In a cross-sectional view of the cornea, five layers can be identified from the outside to the inside (Figure 5-18): (1) the anterior epithelium (stratified squamous nonkeratinizing epithelium), (2) subepithelial basement membrane (Bowman membrane), (3) substantia propria, or stroma, (4) posterior limiting lamina (Descemet membrane), and (5) posterior endothelium (Descemet endothelium).
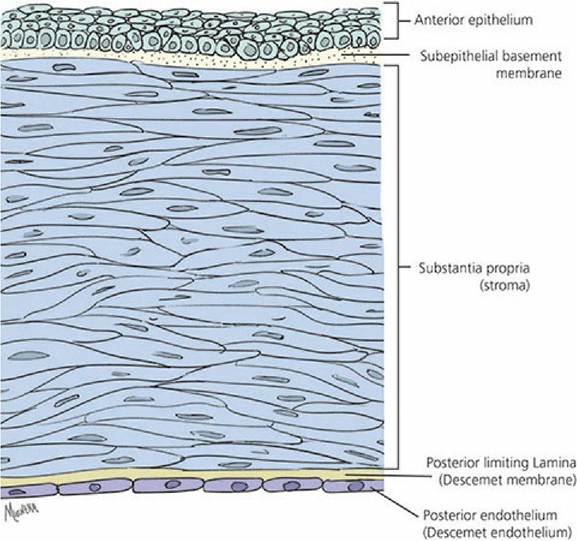
■ FIGURE 5-18 The histologic organization of the cornea (top, anterior surface; bottom, posterior surface).
There is greater light transmission if the surface area ratio of cornea to sclera is increased. Nocturnal animals (nighttime) have relatively larger corneas than diurnal animals (daytime).
About 17% of the eyeball in the dog is cornea (a diurnal animal), whereas about 30% of the eyeball in the cat is cornea (a nocturnal animal).About 90% of the corneal thickness is a result of collagen fibers (called stroma). The collagen fibers have an orderly, laminated arrangement, which is related to the transparent nature of the cornea.
The cornea is avascular (without a blood supply) so that blood vessels do not interfere with the inward transmission of light. The cornea is supplied abundantly with nonmyelinated nerve fibers, bare nerve endings that enter from the limbus and penetrate the outer epithelial layer. The cornea is one of the most sensitive tissues of the body.
Transparency of the cornea depends further on the degree of its hydration; the normal transparent cornea contains less water than it is able to imbibe. Increased uptake of water with a consequent reduction in transparency can occur as a result of damage to either the anterior epithelium or the posterior endothelium or to a reduction of oxygen. If this occurs, a rearrangement of the collagen fibers results, causing the cornea to become cloudy or white. Other causes of corneal cloudiness or whiteness are thinning from increased intraocular tension, disruption by trauma, or replacement by scar tissue.
The Lens and Accommodation
The lens is positioned between the cornea and retina. It is attached by suspensory ligaments (zonular fibers) to the ciliary body, which is a thickened forward ridge of the choroid that circumscribes the eyeball (Figure 5-19). The ciliary body contains three sets of smooth muscle fibers (ciliary muscles), with each set oriented in a different direction. Normal ciliary muscle tone results in normal vision with a lens that is convex and that has a focal distance (distance from the lens to the retina) where the image is focused on the retina. Convex lenses converge light rays. A more convex lens would shorten the focal distance and a less convex lens would lengthen the focal distance.
Accordingly, when nearer objects are viewed, the focal distance is in the back of the retina and the lens must become more convex to shorten the focal distance and bring it to focus on the retina. This is accomplished by increased ciliary muscle contraction. Because the ciliary muscles encircle the eyeball, contraction decreases the tension on the lens ligaments making them more slack (see Figure 5-19). Because of its elastic capsule, the reduced tension on the ligaments allows the lens to assume a more convex configuration and the focal distance is shortened. When more distant objects are viewed, the focal distance is in front of the retina and the lens must become less convex (more divergent) in order to increase the focal distance to the retina. Greater relaxation of the ciliary muscles occurs and more tension is applied to the elastic lens capsule, thereby making the lens less convex, and the focal distance is increased because of the greater divergence. The adjustments needed for near and far objects are known as accommodation. Accommodation among domestic animals seems limited. This is thought to be true because of the sparseness of ciliary muscles, except in the cat. In the cat, the convexity of the lens increases during accommodation for near objects to the extent that it may compress the iris anteriorly to the cornea.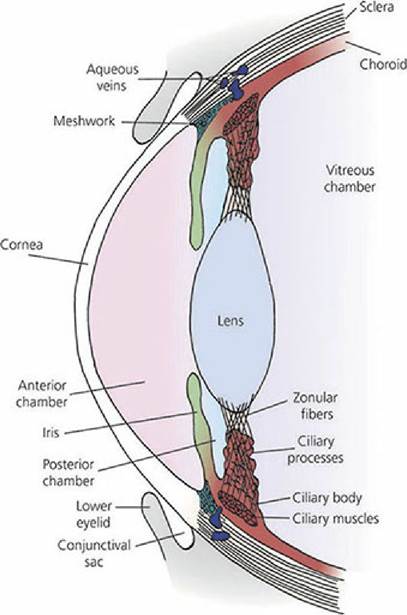
■ FIGURE 5-19 Schematic representation of the relationship in the dog of the ciliary processes with the ciliary body, the zonular fibers (lens ligaments), and the posterior chamber. The ciliary muscles are a part of the ciliary body and are attached to the zonular fibers. The meshwork represents a collection location for aqueous humor that drains to aqueous veins in the sclera. The ciliary muscles encircle the eyeball. When contracted, tension on the lens ligaments is reduced and the lens becomes more convex.
Visual acuity is the extent to which details and forms of objects can be perceived accurately.
A foveal region is an area of the retina where there is high visual acuity and is characterized by a pit (fovea centralis). In primates and birds it contains only cone (color) cells. Domestic mammals lack a foveal region, but do have areas with high visual acuity, called visual streaks.Iris
The amount of light allowed to enter the eye is controlled by the iris, which is the colored part of the eye (see Figure 5-19). The allowed opening, of varying size, is called the pupil. The pupil is horizontal in the domestic herbivores and pig, vertical and elliptic in the cat, and circular in the dog. The iris contains two sets of smooth muscles: (1) circularly arranged fibers, innervated by the parasympathetic division of the autonomic nervous system, and (2) radially arranged fibers, innervated by the sympathetic division. Contraction of the circularly arranged fibers decreases the size of the pupil and allows less light to enter the eye, whereas contraction of the radially arranged fibers increases the size of the pupil and allows more light to enter the eye.
Humors of the Eye The spaces forward from the lens are divided by the iris into two parts. The space behind the iris and forward from the lens is,called the posterior chamber and the space behind the cornea but in front of the iris is called the anterior chamber. Projecting from the ciliary body and into the posterior chamber are structures known as ciliary processes (see Figure 5-19). They present a considerable surface area because of their folded arrangement and are well vascularized. They actively secrete a liquid into the posterior chamber, the aqueous humor (Figure 5-20). The aqueous humor has free communication with the anterior chamber and thus occupies all spaces anterior to the lens. The transparent material behind the lens that occupies most of the volume of the eyeball (the vitreous chamber) is called the vitreous body. It does not have the flow characteristics of a liquid, but rather it is more similar to a gelatinous mass - hence, the name vitreous body is more appropriate than vitreous humor.
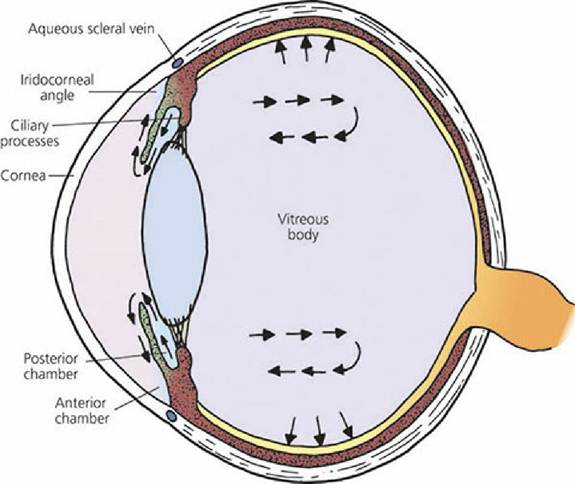
■ FIGURE 5-20 Formation of aqueous humor by ciliary processes and its anterior flow. Arrows in the vitreous body indicate flow by diffusion of aqueous humor through the vitreous body. Some absorption occurs from the vitreous into choroidal vessels.
Aqueous humor can diffuse through the mass of the vitreous body, but only slowly. The principal flow of aqueous humor after its formation is through the pupil into the anterior chamber and to where it is reabsorbed at the iridocorneal angle, which.is the angle formed where the cornea meets the iris (Figure 5-21). Inasmuch as there is a constant formation of aqueous humor, there must also be a constant removal. It enters the corneoscleral meshwork at the iridocorneal angle and is directed to aqueous collecting veins and the scleral venous plexus for its return to blood. A canal of Schlemm does not exist for domestic animals as described for humans. Aqueous humor functions to: (1) provide nutrition to the avascular lens and cornea, (2) remove waste products of metabolism from these structures, and (3) occupy space and maintain a constant distance for the refractive parts. The pressure maintained by the aqueous humor within the eyeball can be measured; it is about 20 mm Hg in the dog. This pressure maintains the normal shape and firmness of the eyeball. If the reabsorption of aqueous humor is impeded, the pressure increases. This situation is recognized clinically as glaucoma and can lead to blindness if left untreated.
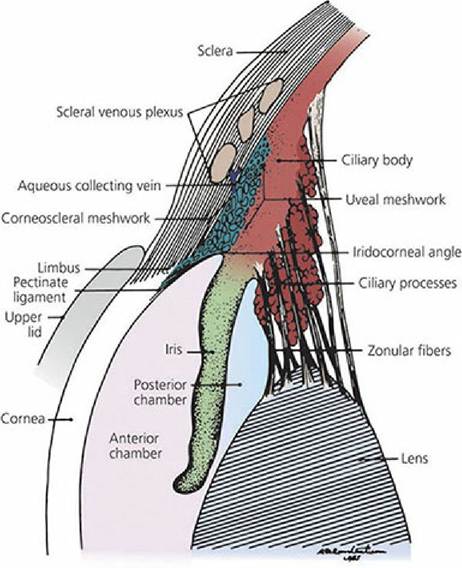
■ FIGURE 5-21 Schematic drawing of the iridocorneal angle region of a dog. This is the location for drainage of aqueous humor. (From Dellmann HD. Textbook of Veterinary Histology. 4th edn. Philadelphia, PA: Lea & Febiger, 1993.)
Retina
The innermost tunic of the eye is the nerve tunic, or neural retina (Figure 5-22). The photoreceptors, the rods and cones, are located near the outer aspect, immediately inward from the pigmented epithelium. Impulse transmission is directed inwardly toward the vitreous. Considerable convergence of impulses from the photoreceptors occurs on two interposed cell layers, of which the innermost cell layer is that of ganglion cells. The unmyelinated axons of the ganglion cell layer arc toward the optic disk, also known as the optic nerve head. Here they become myelinated and turn to form the optic nerve. The intraocular myelinated portion of the nerve forms the optic disk. There are no photoreceptors overlying the disk and hence the optic disk represents a blind spot. It is located ventrolateral to the posterior pole of the eyeball. The posterior pole is the posterior location of the optical axis, which is a line drawn from the center point of the cornea to the center point of the posterior sphere.
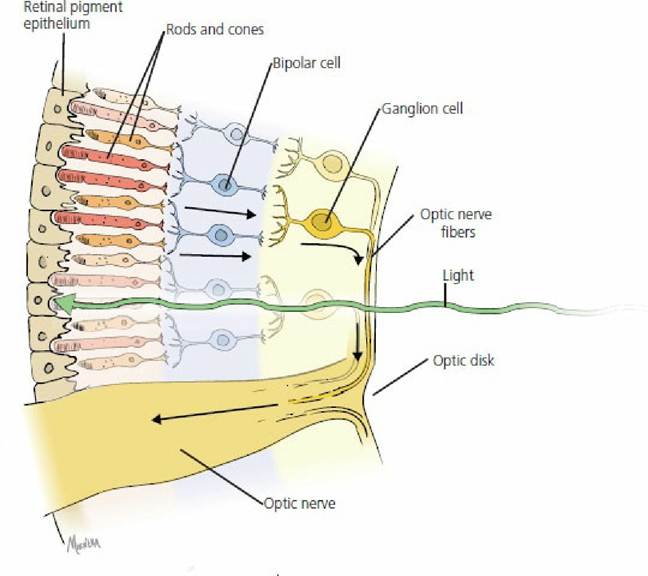
■ FIGURE 5-22 A simplified version of the retina. The arrows indicate the direction of impulse transmission from rods and cones in the outer aspect to the ganglion cells in the inner aspect. Impulse transmission is opposite to the direction of light.
The retinas of domestic mammals contain mostly rods and the retinas of domestic birds contain mostly cones. The rods are the photoreceptors associated with black-and-white vision and the cones are those associated with color vision. The rods are extremely sensitive to light and are used for night vision, whereas cones function best in day vision.
The part of the retina and all associated structures that are visible with the ophthalmoscope are referred to clinically as the ocular fundus. The fundi of several domestic animals are shown in Figure 5-23.
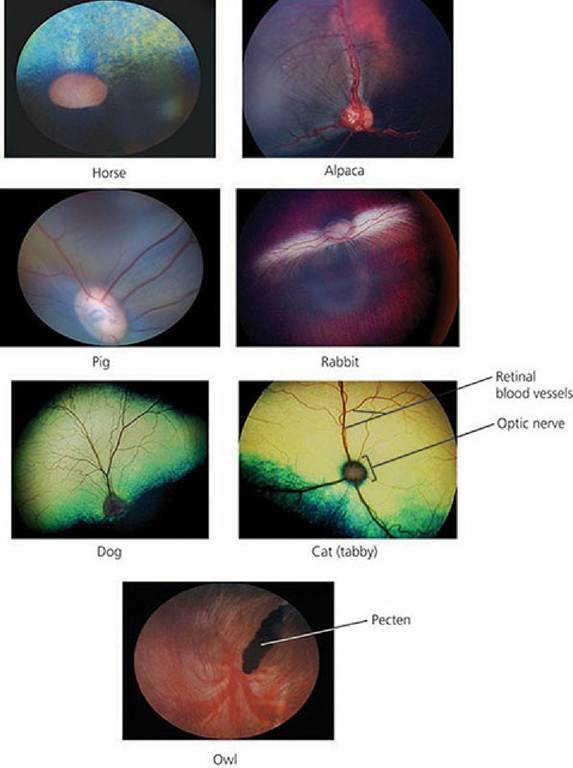
■ FIGURE 5-23 Photographs of the fundi of seven animals as seen by ophthalmoscopy. The diskshaped structures are optic disks shown as an optic nerve head. Pecten, as shown for birds, is responsible for nourishment of the inner eye and retina. (Photographs courtesy of Drs Rachel Allbaugh and Gil Ben-Shlomo, Lloyd Veterinary Medical Center, Ophthalmology Service, Iowa State University.)
Chemistry of Vision
Light that enters the eye stimulates biochemical reactions in the rods and cones. Chemicals in the rods and cones decompose on exposure to light. The chemical in the rods is called rhodopsin and the light-sensitive chemicals in the cones are only slightly different from rhodopsin. The reaction scheme shown in Figure 5-24 is characteristic of the visual cycle.
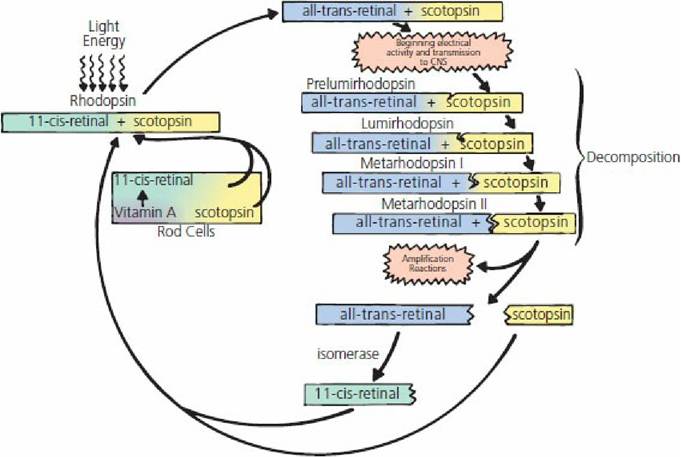
■ FIGURE 5-24 Photochemistry of the visual cycle. Metarhodopsin II, called photoexcited rhodopsin, triggers highly amplified visual excitation. CNS, central nervous system.
Rhodopsin (also known as visual purple) is a light-sensitive pigment in the outer part of the rod that is located in the pigmented epithelium. It is composed of 11-c⅛-retinal (also known as retinene) and scotopsin. Scotopsin is a rod protein and the similar opsin in cones is photopsin. Exposure of rhodopsin to light energy immediately begins its decomposition, in which a number of unstable, short-lived (nanoseconds for prelumirhodopsin and seconds for metarhodopsin II) intermediates are formed. The final one, metarhodopsin II, triggers highly amplified visual excitation and splits into scotopsin and all-trans-retinal. All-trans-retinal is chemically the same as 11-c⅛-retinal but has a different physical structure; it is a straight rather than a curved molecule. Its conversion to 11-c⅛- retinal requires the presence of the retinal enzyme isomerase. All-trans-retinal is converted to 11- c⅛-retinal, which then recombines with scotopsin to reform rhodopsin.
Rod stimulation is believed to occur at the instant that the rhodopsin molecule becomes excited by light. The stimulation resulting from an instantaneous flash of light can persist for about 0.05 to 0.5 s, depending on the intensity of the light. Rapidly successive flashes with alternating intensity become fused to give the appearance of being continuous. This effect is observed when watching motion pictures or television.
There is a relationship between vision and vitamin A. A lack of vitamin A results in inadequate formation of rhodopsin. Night vision requires optimum amounts of rhodopsin and its shortage, because of vitamin A deficiency, is referred to as night blindness.
Adaptation to Varying Light
Dark adaptation refers to an adaptation to relatively dark environments. Because of less light, the concentration of rhodopsin increases, allowing for maximum reaction to the available light. When first entering a dark room, one might be almost unable to see anything, but after dark adaptation, objects can be perceived more readily.
Light adaptation refers to an adaptation to lighter environments. The higher concentration of rhodopsin decomposes because of the abundance of light. The images perceived seem to be overexposed. Normal vision returns when the rhodopsin concentration is balanced with the available light.
Concurrent with the adaptation processes are the visual reflexes, which increase or decrease the diameter of the pupil (see the previous section). Consequently, not only does rhodopsin concentration increase in the dark, but the pupil diameter also increases to allow for maximum light entry. Conversely, rhodopsin concentration decreases in light, and pupil size decreases to minimize light entry.
The tapetum is a light-reflecting layer of cells of the inner choroid, located just outside the retinal- pigmented epithelium (Figure 5-25). Melanin is absent from the pigmented epithelium of the retina where the tapetum is present. The tapetum is not present throughout the choroid and varies in size among those domestic species in which it is present (e.g., cats, dogs, horses, ruminants). The tapetum allows light that has just stimulated the receptor cells to be reflected back on to them so that they receive another stimulation. In this way greater vision is obtained, even with minimal light. The reflected light continues on a forward path through the pupil and out of the eye again. This reflected light is termed eyeshine, which is when eyes glow at night in the presence of light.
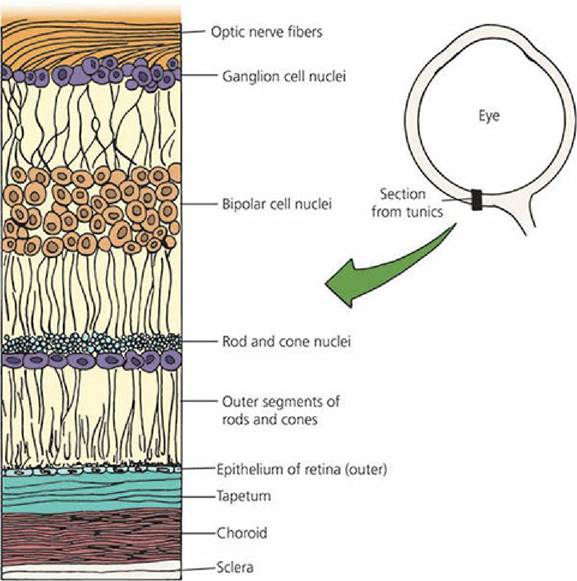
■ FIGURE 5-25 Location of the tapetum relative to the retina. The tapetum is shown as a broad band of cell layers between the choroid and the retina. Melanin is absent from the pigmented epithelium of the retina (outer layer of the retina), where tapetum is present. There is a pigmented layer in the choroid to aid light absorption.
Field of Vision
The field of vision for an animal is the spatial area from which the complete image is formed. The more lateral the placement of the eyes, the larger the field of vision. In fact, some animals might even see everything around them with the exception of objects directly behind their body, which can still be seen with only a slight movement of the head. If the field of vision for each eye overlaps that of the other, a binocular area of vision is formed; conversely, a monocular area of vision is formed if there is no overlap. Binocular vision provides greater depth perception, and this is more pronounced in animals that prey on other animals for food. Greater accuracy of position is necessary before the leap. Such animals characteristically have more forward-placed eyes (Figure 5-26). In contrast, herbivores (plant eaters) have more laterally placed eyes and have a wider field of vision; this gives greater protection while grazing as far as predator observation is concerned (Figure 5-27). In all domestic animals, regardless of how far their eyes are situated laterally, there is some central area of overlap providing a zone of binocular vision.

■ FIGURE 5-26 Field of vision of the cat. The large central binocular area results from the forward position of the eyes. (From Coulter DB, Schmidt GM. Special senses I: Vision. In: Swenson MJ, Reece WO, eds. Dukes’ Physiology of Domestic Animals. 11th edn. Ithaca, NY: Cornell University Press, 1993. Used by permission of the publisher, Cornell University Press.)
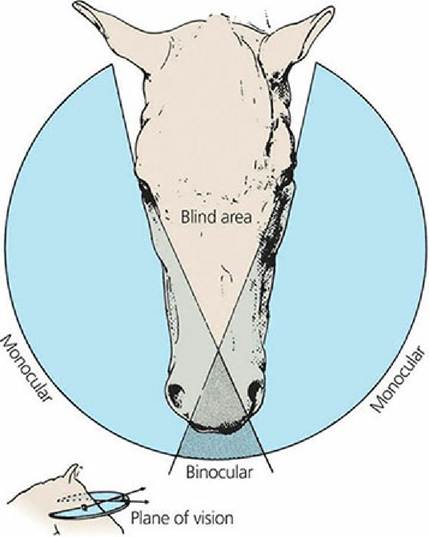
■ FIGURE 5-27 Field of vision of the horse. It is relatively large because of the horse’s more laterally placed eyes. Note the small binocular area. (From Coulter DB, Schmidt GM. Special senses I: Vision. In: Swenson MJ, Reece WO, eds. Dukes’ Physiology of Domestic Animals. 11th edn. Ithaca, NY: Cornell University Press, 1993. Used by permission of the publisher, Cornell University Press.)
The horse has little or no accommodation. When observing more distant objects, the horse increasingly raises its head and lifts up the nose. When observing much closer objects, the horse may arch the neck and rotate the head on one side. In the past, these behaviors were explained as an attempt to make up for the lack of accommodation by using a ramp-shaped retina. This was presumed to provide a longer focal distance for viewing downward than for viewing along the axis of the eye. It was assumed that this retinal feature accounted for the alterations in head position whereby the animal was finding a distance from lens to retina appropriate for focusing the image.
It is now known that the ramp-shaped retina does not exist. Measurements have been made indicating that the horse has a retina that is equidistant from the lens except in the far dorsal and far ventral retina. In these peripheral regions, the retina is nearer to the lens, not farther as proposed for the ramp-shaped retina.
Ganglion cell densities in the horse have been mapped throughout the retina and correlated with maximum visual acuity. Densities of cells are low in the periphery and high in the ventrally placed visual streak, which is a strong narrow region visible in the ventral retina immediately above the optic nerve head. In retinal regions other than the visual streak, acuity in the horse is very low. Acuity is similar at any point along the narrow streak, and the horse can see a narrow, very circumscribed frontal and circular view. Because peripheral acuity is quite low, it would be of little benefit for a horse to use any part of the retina other than the visual streak for direct observation. When the horse lifts its head and points its nose forward to use its binocular field to scan the horizon, its monocular field is lessened and lateral vision becomes more limited (see Figure 5-27). When the animal lowers its head, such that the nose approaches the vertical, the binocular vision is directed toward the ground for grazing and the lateral monocular fields are again in a position to scan the lateral horizon.
The horse can attend to either the frontal field with the head raised or the lateral field with the head lowered. The horse has a frontally placed blind field such that when the nose is drawn in and the face approaches the vertical, the animal is unable to see directly in front. This situation occurs when the horse is being ridden “on the bit” with the neck arched and the nose just in front of the vertical. If a show jumping horse is to see and judge the distance of a fence that it approaches, it must have the ability to raise its head and direct its binocular field forward.
Eyeball Movements and Accessory Structures
Movements of the eyeball are accomplished by skeletal muscles that are innervated by cranial nerves (Figure 5-28). Up-and-down, side-to-side, rotational, and inward (retraction) movements are possible. These muscles also hold the eyeball within its orbit against a pad of fat. Side-to-side movements are made by contraction of a laterally placed muscle; up-and-down movements are made by contraction of a dorsally and ventrally placed muscle; and rotational movements are made by contraction of a dorsal or ventral obliquely placed muscle. The dorsal oblique muscle rotates the top of the eye medially and the ventral oblique muscle rotates the ventral part of the eye medially. Retractor muscles are absent in humans; they seem to provide protection for animals in situations in which protruding eyeballs would be hazardous. Also, retraction of the eyeball causes the third eyelid (nictitating membrane) to slide over the eyeball and spread the tear film.
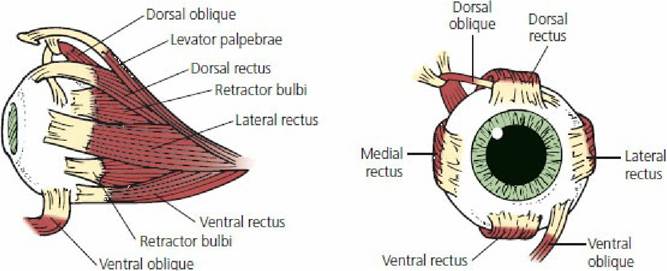
■ FIGURE 5-28 Extrinsic muscles of the eye of the dog. (Adapted from Helper LC. Magrane’s Canine,Ophthalmology. 4th edn. Philadelphia, PA: Lea & Febiger, 1989.)
Conjunctivae
The conjunctivae are the membranes that line the eyelids and that turn back on to the eyeball (see Figure 5-19). The part lining.the eyelid is the palpebral conjunctiva and the part turned back on to the eyeball is the bulbar or ocular conjunctiva. The space between the palpebral conjunctiva and the eyeball forms the conjunctival sac. This is normally minimal and provides a reservoir for the accumulation of tears. It is also used for the application of eye drops and ointments. The conjunctival membrane, because of its superficial location, is useful for examining mucous membrane color. A pink color is considered normal, a blanched appearance indicates lack of blood or anemia, a blue color indicates lack of oxygen, and a yellow color is associated with icterus.
LacrimalApparatus
The lacrimal apparatus is associated with the formation of lacrimal secretions (tears), transport to the conjunctival sac, and drainage to the nasal cavity (Figure 5-29). The lacrimal gland is located in the orbit, dorsal to the eyeball. Short ducts carry the secretion into the upper aspect of the dorsal conjunctival sac. The lacrimal secretion keeps the eyeball moist, provides lubrication, and keeps it clean and free of foreign materials. Ducts (nasolacrimal ducts) in the medial aspect of each eye conduct excess secretion to the nasal cavity, where it is dissipated. If these ducts become occluded, the lacrimal secretions accumulate in the conjunctival sac and overflow on to the face. Glands that secrete a waxy substance, known as meibomian glands, are located along the margin of the eyelids. Their secretion helps to form a dam so that the lacrimal secretion does not ordinarily flow out on to the face.
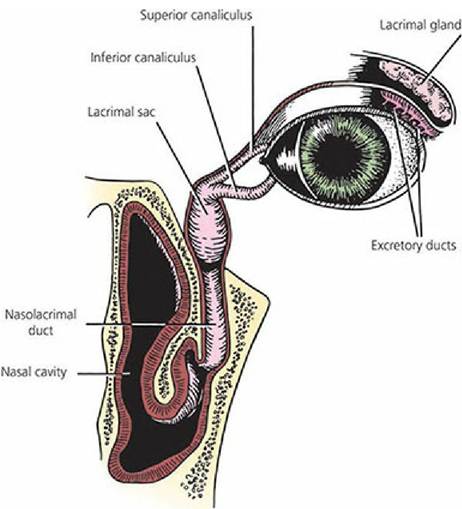
■ FIGURE 5-29 The lacrimal production and drainage system of the eye of the dog. The accessory lacrimal glands are not shown. (From Helper LC. Magrane’s Canine Ophthalmology. 4th edn. Philadelphia, PA: Lea & Febiger, 1989.)
Precorneal Film
The fluid layer on the cornea is known as the precorneal film (also called the tear film). It consists of an innermost layer of mucin, a middle layer of lacrimal secretions (tears), and an outer oil film.
The outer oily layer is formed by meibomian glands and accessory sebaceous glands. It reduces the rate of evaporation of the underlying tear layer and also helps to prevent overflow at the lid margin.
The middle layer of fluid is composed of lacrimal secretions, which wet the cornea and reduce evaporation from the eye. It is formed by the lacrimal glands and the accessory lacrimal glands (such as those associated with the third eyelid).
The inner mucinoid layer is formed by the goblet cells of the conjunctiva. In addition to mucin, it contains a high concentration of lysozyme (an enzyme), which can digest bacterial cell walls.
Lysozyme is present in most animal tissues and secretions, but it only is found in sufficiently high concentrations to be bactericidal in white blood cells, nasal secretions, and tears. In addition to lysozyme, a gamma globulin protein fraction also contributes to the antibacterial property of tears. Wettability (favorable surface tension between cornea and tears) is provided for by the mucin of the inner layer.
The tear film is reformed each time the eyelids close or when the third eyelid is swept across the eye by eyeball retraction.
Third Eyelid
The third eyelid arises as a fold from the ventromedial aspect of the conjunctiva (the visible part shown as membrane nictitans in Figure 5-16). It is well developed in the dog, is highly mobile, and is large enough to cover the entire anterior face of the cornea. The third eyelid is reinforced by a T- shaped cartilage surrounded at its base by a gland that contributes to the tear film (Figure 5-30). Pigs and cattle also have a second, deeper gland. The third eyelid becomes prominent when all of the muscles of the eyeball are caused to contract, as in some clinical situations such as strychnine poisoning. The contraction retracts the eyeball and puts pressure on the cartilaginous plate, pushing it forward. Lymph nodules are also present on the underside of the third eyelid. When they become inflamed in the dog, they often protrude; the condition is referred to as “cherry eye.”
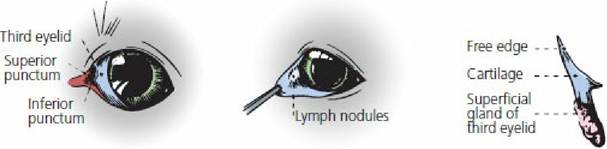
■ FIGURE 5-30 Third eyelid in the dog. (From Evans HE, deLahunta A. A Guide to the Dissection of the Dog. 8th edn. St Louis, MO: Elsevier, 2017.)
■