HEARING AND EQUILIBRIUM
1. Be able to follow the motion initiated by a sound wave from the tympanic membrane through the cochlear window (round window).
2. What is the function of the two striated muscles located in the middle ear?
3.
What reflex is inherent to the function of the middle ear muscles?4. What are the respective functions of the vestibular and cochlear portions of the inner ear?
5. Why is the cochlea coiled?
6. Differentiate between membranous labyrinth and osseous labyrinth and their respective fluids.
7. How are the cristae (located in the semicircular canals) stimulated?
8. How are the macula receptors stimulated?
9. What are the divisions of the cochlea brought about by extension of the membranous labyrinth into the cochlea?
0. Which cochlear division contains the organ of Corti?
11. What function is served by the organs of Corti?
2. Summarize sound reception (which relates to item 1).
The ear has components associated with the special sense of hearing and those involved with positional equilibrium. Sound waves are directed to the receptors for hearing in the inner ear via the external ear and middle ear. The inner ear has the receptors not only for hearing but also for equilibrium.
External Ear
The external ear (Figure 5-5) consists of the outer visible part (the pinna) and ear canal that extends from the pinna into the substance of the skull (external acoustic meatus) to the middle ear (tympanic cavity). The pinna in most animals consists of a funnel-shaped cartilage that is lined on the outside with skin having a generous amount of hair and on the inside with relatively hairless skin. Varying degrees of muscle attachment lend mobility to the pinna, which is helpful in localizing and picking up sounds. The funnel-shaped cartilage concentrates sound waves and directs them through the ear canal toward the tympanic membrane, which separates the middle ear from the external ear.
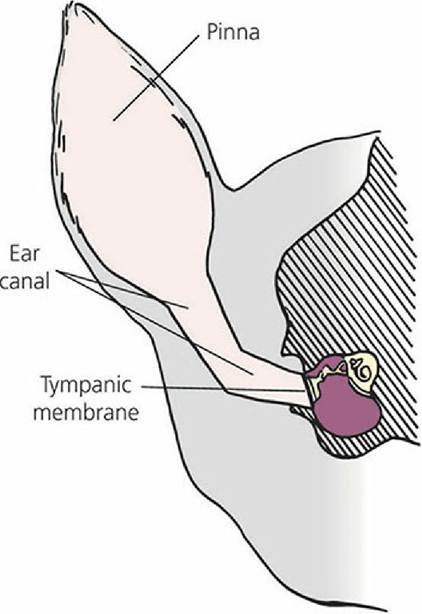
■ FIGURE 5-5 Transverse section through the dog head.
The external ear (pinna and ear canal) provide for transfer of sound waves to the tympanic membrane. The ear canal in the dog has a vertical and an oblique component.Middle Ear
The middle ear and inner ear are shown in Figure 5-6. The middle ear is separated from the inner ear by membranes that close the vestibular (oval) window and cochlear (round) window. The middle ear communicates with the pharynx by way of the auditory tube (often called the eustachian tube). The auditory tube allows for equalization of pressures between an otherwise closed cavity and the outside. Within the middle ear a mechanical linkage is provided between the tympanic membrane and the membrane closing the vestibular window by three auditory ossicles (bones). From lateral to medial they are the malleus, incus, and stapes, or, more commonly, the hammer, anvil, and stirrup. Amplification of sound waves is provided by leverage of the ossicles and by the greater surface area of the tympanic membrane, which transmits sound waves to the smaller surface area of the vestibular window. Excessively loud noises are damped by two skeletal muscles in the middle ear, the tensor tympani and the stapedius muscle (Figure 5-7). Muscle spindles within these muscles respond to muscle stretch by initiating a reflex that causes the muscles to contract. The tensor tympani muscle is attached to the malleus and its contraction tenses the tympanic membrane, thus limiting its movement. The stapedius muscle (the smallest skeletal muscle in the body) is attached to the stapes. Its contraction tenses the stapes to reduce its movement. The degree of stretch is determined by the intensity (loudness) of the sound wave. Loud noises are damped because of excessive muscle stretch and subsequent reflex muscle contraction, which prevents excessive movement of the ossicles.
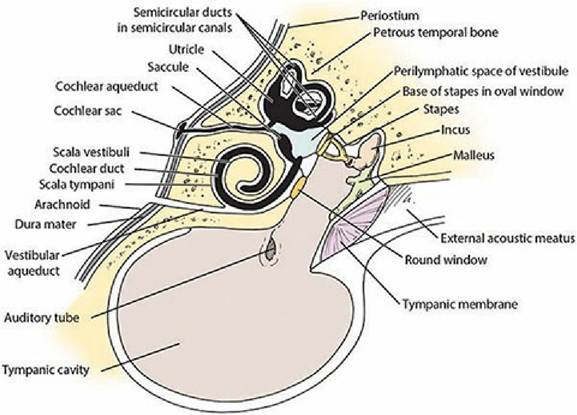
■ FIGURE 5-6 A schematic of the middle ear and inner ear that features the membranous labyrinth (black), which contains endolymph within its ducts, within the osseous labyrinth, which contains perilymph within the semicircular canals and vestibule.
The vestibular aqueduct communicates with the subarachnoid space and its cerebrospinal fluid. (Adapted from Getty R, Foust HL, Presley ET, Miller ME. Macroscopic anatomy of the ear of the dog. Am J Vet Res, 1956; 17: 366.)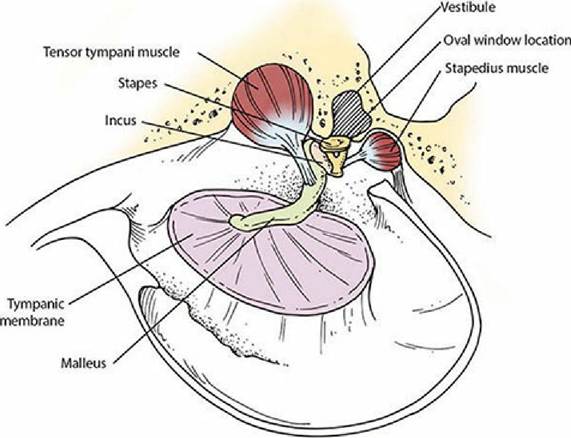
■ FIGURE 5-7 Inside view of the middle ear. The malleus is attached to the tympanic membrane and the stapes is attached to the vestibular (oval) window. Muscle spindles in the tensor tympani and stapedius muscles initiate the stretch reflex in response to loud noises. Contraction of these muscles, respectively, tenses the tympanic membrane, limiting its movement, and reduces movement of the stapes.
Inner Ear
The inner ear can be divided into two parts according to function: (1) the vestibular portion, which is sensory for position and equilibrium, and (2) the cochlear portion, which is sensory for sound (Figure 5-8). The cochlear portion receives the cochlear nerve, a branch of the vestibulocochlear nerve (cranial nerve VIII), and the vestibular portion receives the vestibular nerve branch of the same cranial nerve (VIII). The inner ear is contained within a bony excavation known as the osseous labyrinth (Figure 5-9). Labyrinth refers to an intricate combination of passages. Because the cochlea is coiled, it can occupy limited space. An uncoiled cochlea would project into the brain.
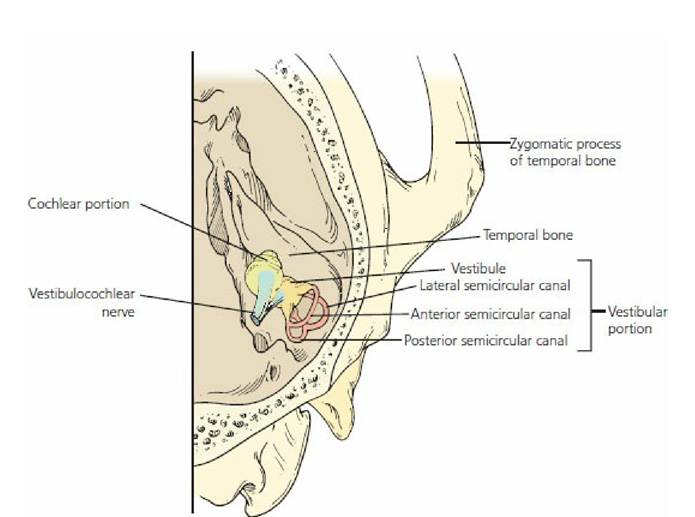
■ FIGURE 5-8 Right inner ear (viewed from above showing its orientation with the skull). (Adapted from Getty R, Foust HL, Presley ET, Miller ME. Macroscopic anatomy of the ear of the dog. Am J Vet Res. 1956; 17: 369.)
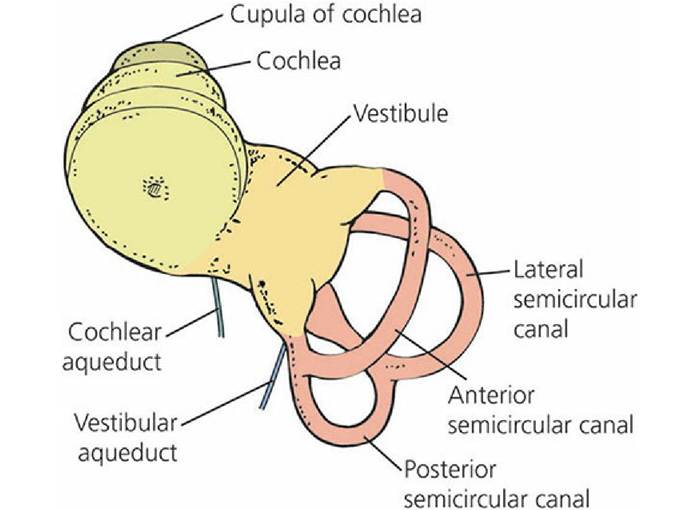
■ FIGURE 5-9 Drawing of a latex cast of the inner ear of the dog, occupying the osseous labyrinth. The membranous labyrinth would occupy the space filled by the latex cast.
(Adapted from Getty R, Foust HL, Presley ET, Miller ME. Macroscopic anatomy of the ear of the dog. Am J Vet Res. 1956; 17: 370.)Vestibular Structure and Function
The vestibular portion is housed in the parts of the osseous labyrinth known as the vestibule and three semicircular canals (anterior, lateral, and posterior). Each semicircular canal leaves and returns to the vestibule (see Figure 5-9). In addition, each semicircular canal is arranged so that it is in a different geometric plane from the others (at right angles to each other). The cochlear portion is housed mostly in.the cochlear portion of the osseous labyrinth that departs from the vestibule.
Within the osseous labyrinth is a membranous labyrinth, which is a completely closed connective tissue structure (Figure 5-10). It contains a fluid known as endolymph (a fluid high in potassium and low in sodium) that is secreted by the vascular tissue on the outer wall of the scala media. Outside the membranous labyrinth and within the osseous labyrinth is another fluid known as perilymph. By way of the vestibular aqueduct (connection to the subarachnoid space), perilymph circulates freely with cerebrospinal fluid and shares a similar composition (see Figure 5-6). Because of the meningeal connection, a potential exists for inner ear infections to ascend to the meninges and produce meningitis (inflammation of the meninges). Within the vestibular portion, the membranous labyrinth also includes three semicircular ducts and two sacs within the vestibule known as the utricle and saccule. Both ends of each semicircular canal open into the utricle, and the utricle communicates with the saccule (see Figure 5-10). The saccule has two other communications, a major one with the membranous labyrinth of the cochlea and another with the cochlear aqueduct, which leads to the cochlear sac that lies between the layers of the meninges (subdural) (see Figure 5-6). The cochlear sac is a site for the active absorption of endolymph.
Also, neutrophils and macrophages from the surrounding connective tissue can cross the epithelium to phagocytize cellular debris and other particulates that may accumulate in the lumen of the cochlear sac.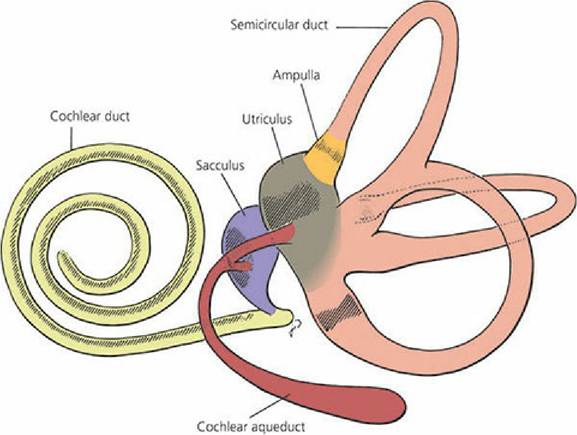
■ FIGURE 5-10 Membranous labyrinth. Hatched regions indicate the sites of neuroepithelium, including the spiral organ of the cochlear duct, the maculae of the utriculus (utricle) and sacculus (saccule), and the crista ampullares of the semicircular ducts. (From Banks WJ. Applied Veterinary Histology. 2nd edn. Baltimore, MD: Lippincott Williams & Wilkins, 1986.)
As each membranous labyrinth occupying the semicircular canals leaves the utricle, a dilated portion is noted - the ampulla. Each of the three ampullae contains sensory receptors for equilibrium known as a crista (Figure 5-11). The utricle and saccule each contain a sensory receptor area known as a macula (Figure 5-12). In both cases, the receptive regions are patches of epithelium containing hair cells, and the vestibular impulses that the hair cells generate contribute to an overall sense of orientation and balance.
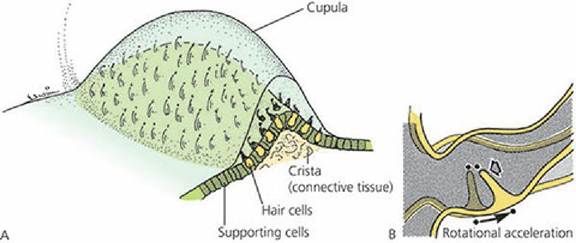
■ FIGURE 5-11 A. General structure of the ampullary crista in a semicircular duct. B. Ampullary
hair cells responding to deflection of the cupula. (From Cormack DH. Ham’s Histology. 9th edn. Philadelphia, PA: JB Lippincott Company, 1987.)
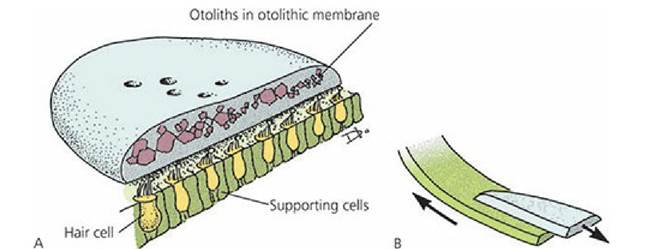
■ FIGURE 5-12 A. General structure of the utricular and saccular maculae. B. Macular hair cells respond to movement of otolithic membranes. (From Cormack DH. Ham’s Histology. 9th edn. Philadelphia, PA: JB Lippincott Company, 1987.)
The maculae in the utricle and saccule (see Figure 5-12) have receptor hair cells that are embedded in an otolithic membrane consisting of a gelatinous material covered by a glycoprotein studded with calcium carbonate crystals called otoliths (otoconia), which are relatively heavy.
The utricle receptors lie in a horizontal plane and those of the saccule lie in a vertical plane. Because of the pull of gravity, and depending on the position of the head, the heavy otoliths on one or both of the maculae can apply shearing stresses to the hair cells. This force registers the position of the head. Because of the weight of the otoliths, sufficient inertia is provided to the,otolithic membrane for the maculae to also sense linear acceleration or deceleration of the head.Rotational acceleration or deceleration involving the head is detected in any given plane by the ampullary crista of the corresponding semicircular duct. Hair cells of the cristae are stimulated when the head is moved because the hair cells are mechanically moved through the endolymph, which does not move as a result of inertia. When the head stops, the endolymph is finally moved; this stimulates the hair cells in an opposite direction and further accommodation is inhibited.
Cochlear Structure and Function
The extension of the membranous labyrinth into the cochlea is known as the cochlear duct or, more commonly, the scala media. This occupies a central position within the cochlea extending from one side to the other and dividing the cochlea into a part above the scala media (scala vestibuli) and a part below (scala tympani),(Figure 5-13). These latter divisions do not communicate with each other except for a small opening at the apex or tip of the cochlea, known as the helicotrema. Along the length of the scala media are a large number of structures, each individually called an organ of Corti (see Figure 5-13). These structures convert sound waves to nerve impulses, which in turn are transmitted to the cerebral cortex to provide the sensation of hearing. The nerve entries for these structures and for the larger basilar cells are arranged so that the thicker base is called the basilar membrane. The location of a particular organ of Corti within the scala media, from the base (near the middle ear) to the apex of the cochlea, with its individual innervation, determines the frequency of the sound wave perceived (Figure 5-14).
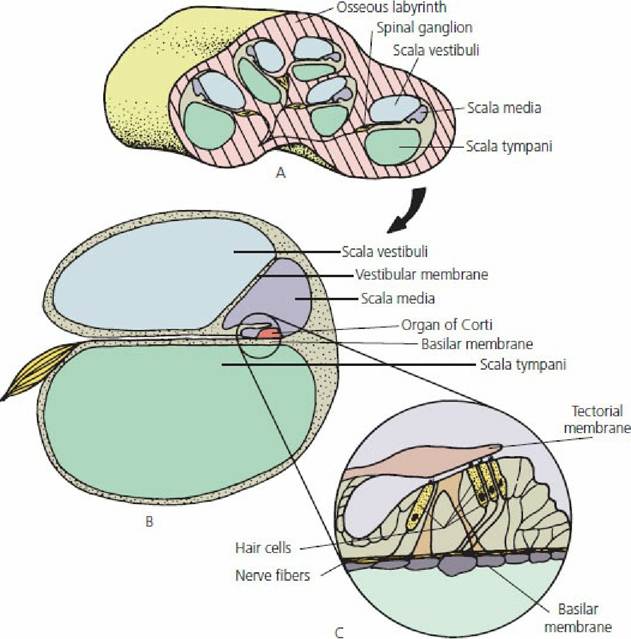
■ FIGURE 5-13 The cochlear portion of the inner ear. A. Cross-section through the cochlea, illustrating its coiled nature. B. Schematic representation of a section through one of the turns of the cochlea. C. Details of an organ of Corti.
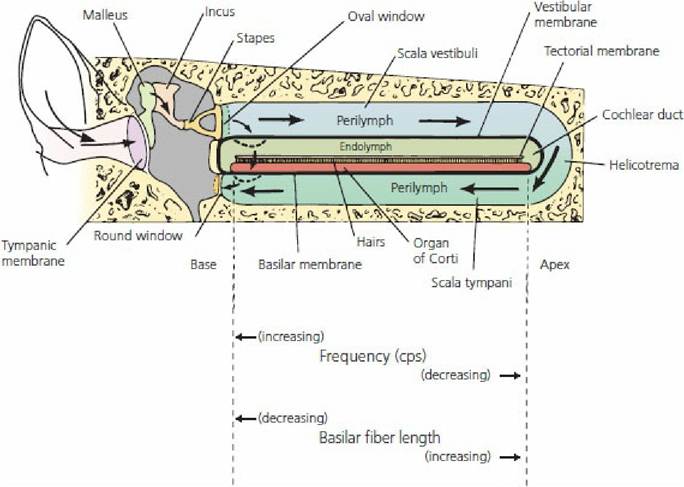
■ FIGURE 5-14 Transmission of pressure waves in the cochlea. When the movement of the stapes is slow, pressure waves are transmitted through the perilymph with no movement of the basilar membrane. With greater movement of the stapes (higher frequency), pressure waves are directed through the endolymph with movement of the basilar membrane, and hence sound is perceived. High- and low-frequency sounds are related to regions of the basilar membrane where different sound wave frequencies can cause displacement. (Adapted from Spence AP, Mason EB. Human Anatomy and Physiology. 4th edn. St Paul, MN: West.Publishing Co., 1992.)
Sound waves of different frequencies have different transmission patterns from the base to the apex. A weak sound wave (of any frequency) at the base strengthens when it reaches the portion of the basilar membrane that has a natural resonant frequency equal to its own. At this point, the basilar membrane can vibrate easily; the sound wave energy is dissipated and does not travel the remaining distance along.the basilar membrane. Therefore, a high-frequency sound wave travels a short distance along the basilar membrane, where it reaches its resonant point and dies out. A low- frequency sound wave travels a longer distance and a similar phenomenon occurs. All frequencies between high and low are represented at separate points on the basilar membrane between the base and apex of the cochlea.
An organ of Corti is composed primarily of sensory receptors called hair cells that have hairs projecting toward the tectorial membrane (see Figure 5-13). Displacement of the hair cell cilia against the tectorial membrane caused by oscillations of the basilar membrane (resulting from dissipation of sound waves) causes the hair cells to depolarize and create a nerve impulse, which is transmitted to the auditory cerebral cortex by way of the vestibulocochlear nerve.
Summary of Sound Reception
The structures traversed and actions initiated for a sound wave to be heard may be summarized as follows (Figure 5-15):
1. The sound wave is directed into the external auditory meatus by the pinna.
2. The sound wave strikes the tympanic membrane (eardrum) and sets it in motion.
3. The motion of the eardrum is transmitted through the middle ear by the auditory ossicles to the vestibular (oval) window.
4. The vestibular window is set in motion and displaces perilymph in the vestibule of the inner ear.
5. The perilymph (a liquid) is incompressible and thus transmits the sound wave through the scala vestibuli of the cochlea.
6. An organ of Corti at a distance from the base characteristic of the approaching sound wave is stimulated when a sound wave is transmitted to the scala media and from there to the scala tympani.
7. The movement of the liquid in the scala tympani is finally compensated for by an outward movement of the cochlear (round) window into the cavity of the middle ear.
8. The stimulation of hair cells in an organ of Corti initiates a nerve impulse that is transmitted by the cochlear branch of the vestibulocochlear nerve to the brain.
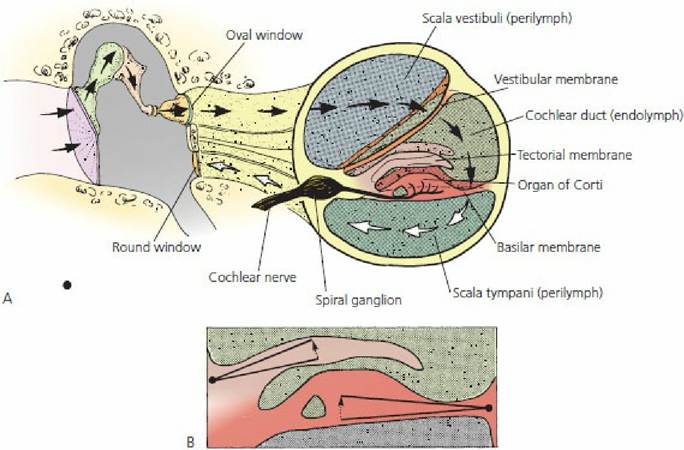
■ FIGURE 5-15 A. Schematic representation of the pathway of sound waves that enter the ear. B. Hair cells in the organ of Corti respond to shearing stresses generated by independent pivoting of the tectorial and basilar membranes on separate axes. (From Cormack DH. Essential Histology. 2nd edn. Baltimore, MD:.Lippincott Williams & Wilkins, 2001.)
The range of frequencies through which sound can be perceived varies among species. In humans, the limit seems to be between 20 and 20,000 cycles per second (cps). Dogs can perceive frequencies up to about 50,000 cps, which is the basis for dog whistles. These emit a high-frequency sound that is not perceived by humans, but the dog responds because its organ of Corti is stimulated.
■