Volatile Fatty Acids, Representing 60% to 80% of the Energy Needs of the Animal, Are Absorbed Directly from the Forestomach Epithelium
VFAs are bacterial waste products and, if allowed to accumulate, will suppress fermentation. Furthermore, the VFAs are extremely important energy substrates for the host» supplying 60% to 80% of the dietary energy to ruminants with most types of diets.
Therefore the presence of an efficient and high-capacity mechanism for VFA absorption is important to both digestion and host metabolism. The forestomach epithelium supplies such a system, absorbing almost all the VFAs, with only small amounts escaping to the lower digestive tract. In addition, the absorptive process helps maintain rumen pH by removing acid from the forestomach ingesta and contributing bicarbonate in the process.The epithelium responsible for this tremendous absorption is structurally much different from other absorptive epithelia of the GI system. However, the nature of the rumen epithelium may give it functional characteristics similar to those of the absorptive epithelium of the small intestine and colon. The forestomach surface is of the stratified squamous type and, as with the stratified squamous epithelium of the skin and other surfaces, consists of several layers of cells of varying maturity. The deepest layer is the stratum basale, from
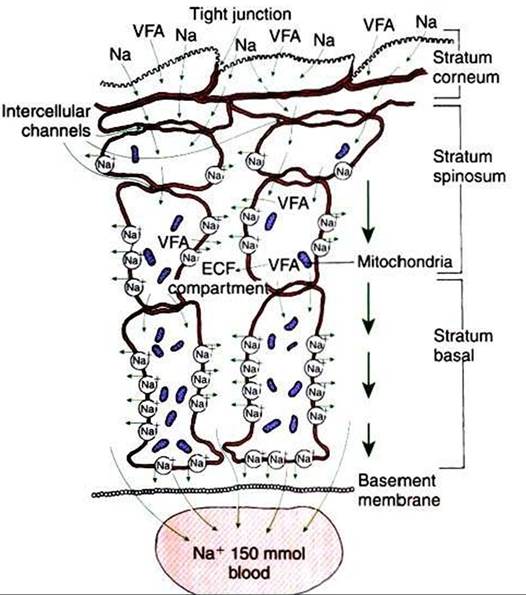
FIGURE 31-11 The stratified squamous epithelium of the rumen, although anatomically much different, shares functional similarities with the columnar epithelium of the intestine. Note the tight junctions of the cells of the stratum corneum and the lateral space-like compartment between adjacent cells of the stratum spinosum and stratum basale. Although the cells of the stratum spinosum are metabolically inactive, the intercellular channels allow the metabolic actions of the stratum basale to be reflected in the more superficial layers.
VFAf Volatile fatty acid; Naf sodium; ECFf extracellular fluid. {Modified from Steven DH, Marshall AB: Organization of the rumen epithelium. In PhiIIipsonAT, editor: Physiology of digestion and metabolism in the ruminant, Newcastle upon Tyne, UK, 1970, Oriel Press.)which cells divide and migrate into the stratum spinosum. Cells of the stratum spinosum begin the process of keratin- ization and continue into the stratum granulosum, which is covered by the outermost and most keratinized layer, the stratum corneum. Although the forestomach epithelium seems completely different from the columnar epithelium of the small intestine, an interesting similarity between forestomach and intestinal epithelia is noted when the cellular attachments and intercellular spaces of the forestomach are examined (Figure 31-11).
The cells of the stratum granulosum are tightly joined by junctions that may functionally resemble the tight junctions of the enterocytes (see Chapter 30). Deeper in the epithelium, the cells of the stratum spinosum and stratum basale are separated by intercellular spaces that increase in size as the basement membrane is approached. These intercellular spaces are reminiscent of the lateral spaces of columnar absorptive epithelia. If these observations are combined with the existence of the intercellular bridges that characterize the forestomach epithelium, an interesting analog}' to columnar absorptive epithelia can be constructed. VFAs, electrolytes, and water apparently are initially absorbed through the stratum corneum and passed cell to cell by way of intercellular bridges to the cells of the stratum spinosum and stratum basale, from which the absorbed substances are passed into the intercellular spaces before entering the capillaries.
This arrangement of the forestomach epithelium is very similar to the three-compartment characteristics of the columnar absorptive epithelia, with solutes passing from lumen to cell to lateral spaces.
Although the keratinized cells of the stratum corneum do not appear to retain adequate metabolic machinery (e.g., mitochondria) to maintain appropriate gradients for diffusion, the cells of the stratum spinosum and stratum basale are metabolically active. Because of the intercellular bridges, absorbed solute can be transferred directly from the outer keratinized cells to the deeper, more metabolically active cells. Thus the metabolic activity deep in the epithelium appears to maintain conditions for absorption at the epithelial surface.The molecular mechanism of VFA absorption is incompletely understood but seems to involve local alterations in pH near the absorptive surface. Differences in pH can have an important influence on VFA absorption because of shifts in the dissociation state of the VFA molecules. The pKa of the VFA is approximately 4.8, well below the normal pH of the rumen; thus most of the VFAs exist in the rumen in the dissociated, or ionic, form. However, sodium-hydrogen ion exchange by the epithelial cells may decrease the local pH at the absorptive surface. Such a drop in pH would lead to a shift in the VFA from the ionic to the free-acid state. Cell membranes are permeable to VFA free acids, and absorption proceeds because of the concentration gradient between the lumen and cells. The high CO2 tension in the rumen, caused by the production of fermentation gases, may also enhance the conversion of VFA to the free-acid state. As shown in Figure 31-12,
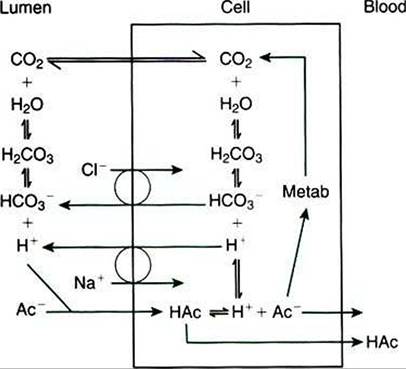
FIGURE 31-12 - VFA absorption is promoted by the conversion of VFA anions (Ac~) to free acids (HAc) in the microenvironment near the epithelial surface.This diagram illustrates two proposed means, one intracellular and one extracellular, by which hydrogen ions could be locally generated to effect the formation of VFA free acids; both mechanisms could exist simultaneously. (From Stevens CE, Argenzio RA, Roberts MC: Comparative physiology of mammalian colon and suggestions for animal models of human disorders, Clin Gastroenterol 15:763, 1986.)
when one VFA molecule is absorbed, one molecule of bicarbonate (HCO3") is generated in the lumen; thus VFA absorption helps buffer rumen pH both by generating base and by removing acid.
All the VFAs appear to be absorbed by the same mechanism, but they are handled differently within the epithelial cells. Some acetate seems to be completely oxidized within the cells, with the remainder absorbed unchanged. Most propionate is absorbed, but a small portion is converted to lactate by the epithelial cells. Butyrate is modified extensively, and essentially all molecules are changed to β-hydroxybutyrate before absorption, β- Hydroxybutyrate is an important metabolite known as a ketone body. Ketone bodies are metabolites that frequently have special medical significance (see Chapter 32). In ruminants the rumen itself is a significant source of ketone bodies. In monogastric animals, however, ketone bodies arise exclusively from the partial oxidation of long-chain fatty acids.
The rumen epithelium is arranged in papillae, fingerlike projections that increase the absorptive surface area. Although they serve the same area-expanding function as the villi of the small intestine, papillae are much larger and easily visible to the unaided eye. The size and shape of the papillae are quite dynamic and responsive to changes in diet. Papillary growth is stimulated by VFAs, especially butyrate and propionate. Diets with high digestibility result in high rumen VFA concentrations, which stimulate the growth of long papillae. In contrast, animals receiving little feed or diets of low digestibility have short rumen papillae. It is important to adapt ruminants gradually when changing them from diets of low digestibility to high digestibility; this may allow time for sufficient adjustment of papillary size so that VFA absorption will match VFA production.