Anatomy
The liver is one of the largest organs in the body. It weighs approximately 3% of the total body weight in adult dogs and cats. In very young growing animals, it accounts for 5% of the body weight.
The shape of the liver in dogs and cats is different from that of most other mammals such as cattle, horses, and humans. The domesticated carnivores have deeply incised lobes, which are individually recognizable, whereas they are largely fused in other species. This may be due to the concave shape of the diaphragm in the dog and cat, which leads to relatively large excursions during in- and expiration, so that the liver needs to be able to fold and unfold while following the diaphragm. The deeply separated lobes, which connect at the hilus of the liver, make it relatively easy to resect an entire liver lobe in dogs and cats. By far the largest liver lobe is the left lateral lobe, which encompasses 30-40% of the entire liver mass. This large lobe is therefore the easiest one to approach for the collection of liver biopsies. The peripheral part of the left lateral lobe is far away from critical structures such as large veins, arteries, or bile ducts. From a dorsoventral perspective, the position of the liver is somewhat turned to the right. The gall bladder, large bile ducts, and blood vessels of the liver enter the organ at the hilus in the right upper abdominal quadrant. The liver follows the contours of the diaphragm and the abdominal section of the rib cage.In dogs, the liver lies entirely within the rib cage and, therefore, is normally not palpable. Depending on the shape of the rib cage, the liver lies more or less hidden within it. In brachycephalic dogs, which have a wide rib cage and a more flattened diaphragm, any enlargement will result in the liver being palpable. In contrast, in racing hounds with a deep rib cage, the liver may only become palpable behind the last rib when it is severely enlarged.
In healthy cats, the ventral border of the liver can often be palpated and the feline liver is always palpable when enlarged. Since feline hepatobiliary diseases are usually associated with hepatomegaly, abdominal palpation is often informative in cats with liver disease.The stomach is in contact with the visceral surface of the liver. As a result, changes in size or shape of the liver cause displacement of the stomach. This is one possible explanation why vomiting is often a major clinical sign in patients with liver disease.
The peritoneal capsule of the liver forms ligaments, which fixate the organ in its position while maintaining a degree of movability. The falciform ligament is located in the midline and attaches the liver to the ventral abdominal wall and also contains the abdominal retroperitoneal fat mass. The other fixation points of the liver are the left and right triangular ligaments and the short coronary ligament, which connect the liver with the diaphragm laterally and in the midline, respectively. Finally, the right hepatorenal ligament connects the liver with the right kidney.
7.1.1 Biliarysystem
One of the main functions of the liver is the detoxification or catabolism of many exogenous and endogenous compounds. Many compounds undergo biotransformation in the hepatocytes. Larger molecules (i.e., molecular weight > 300), such as steroid hormones, bilirubin, bile salts, and many toxins are excreted by the biliary system. In contrast, smaller exogenous or endogenous molecules (i.e., urea or allantoin) are excreted via the urine.
The biliary system is a branched tree, which serves as a collection system for bile from each individual hepatic cell. In each hepatic lobule, the hepatocytes are organized in liver cell plates, which radiate around a central vein. These hepatocyte plates connect the central vein with several portal areas. Blood flows from the portal areas to the central vein, while bile flows from the central areas to the portal areas and the common bile duct.
An alternative view (formulated by Rappaport) sees the portal tract in the center of the hepatic lobule and the region supplied by blood vessels originating in this portal tract of the lobule. According to this view, the central veins lie in the periphery of the lobule and are called terminal hepatic veins (THV). However, there is a tendency to return back to using the old nomenclature (central vein), but the terms central vein and THV are often used interchangeably.The lateral membranes connecting adjacent liver cells contain a specialized region, the canalicular membrane. The space between the canalicular membranes of adjacent cells is sealed by tight junctions and forms the smallest branches of the biliary system, the canaliculi. This excretory region of the hepatocyte membrane constitutes 15% of the hepatocellular surface. Most molecules are excreted across this membrane by their own specialized transporters, which can actively excrete against a very high concentration gradient. Thus, the canalicular membrane is a highly specialized section of the hepatocellular membrane.
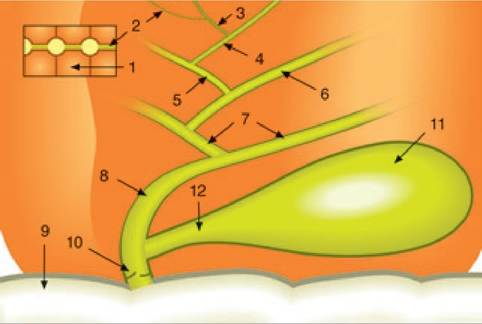
Figure 7.1:
Bile flow. This figure shows a schematic representation of bile flow in dogs and cats. Bile is produced by the transport of a large number of molecules across the canalicular membranes of the hepatocytes (1) into the canaliculi (2). The canaliculi drain into the canals of Hering (3), which drain into the intrahepatic bile ductules (4). These then drain into the interlobular ducts (5), septal ducts (6), and hilar ducts (7), which collect all the bile from one liver lobe. The hilar ducts merge to form the common bile duct (ductus choledochus; 8), which opens into the duodenum (9) on Vater's papilla and is protected against the reflux of duodenal contents by the muscular sphincter of Oddi (10). The gall bladder (11) is connected to the common bile duct by the cystic duct (12).
Bile is excreted by hepatocytes into the canaliculi, which then transport the bile to the portal tracts where they enter the canals of Hering (Figure 7.1).
These short connecting ducts are in part lined by hepatocytes, in part by cuboidal cholan- giocytes.2 Some of the cells of the canal of Hering are the stem cells of the liver, which can differentiate into hepatocytes as well as cholangiocytes, and thus function as stem cells for all the main cell types of the liver. These hepatic progenitor or stem cells are also called oval cells. All hepatocytes have an enormous capacity to divide and thus allow for regeneration of the liver mass after loss due to damage. Therefore, all hepatocytes have stem-cell-like characteristics. However, many biliary epithelial cells can only be replenished by oval stem cells.The canals of Hering drain into the smallest branches of the intrahepatic bile ducts, which are completely lined by cholan- giocytes. From the smallest bile ductules, the bile flows to the interlobular ducts, the septal ducts, and via the hilar ducts of each liver lobe into the common bile duct (ductus choledo- chus). The ductus choledochus is only 2-3 mm in diameter in dogs and cats, and may be difficult to find with ultrasonography. However, in patients with a common bile duct obstruction, dilation of the common bile duct is easily visible ultrasonographically and serves as the hallmark feature of common bile duct obstruction. The common bile duct runs to the duodenum and enters the major duodenal papilla some 3-6 cm caudal to the pylorus, depending on the size of the animal. The junction of the common bile duct and the duodenum, Vater’s papilla, is located close to the head of the pancreas and is surrounded by the muscular sphincter of Oddi. In cats, the pancreatic duct and the common bile duct merge and enter the duodenum together on the major duodenal papilla. This may explain why in cats concurrent pancreatitis and cholangiohep- atitis /cholangitis occur quite frequently. In contrast to cats, the bile duct and the pancreatic duct usually end separately in dogs. The gallbladder is connected with the common bile duct via the short cystic duct.
The gall bladder provides the main storage facility for bile and concentrates the bile approximately tenfold. Bile is also concentrated in the larger bile ducts. In healthy dogs and cats, a completely filled gallbladder contains around 1 ml of bile/kg body weight. Less than half of the bile produced is stored and concentrated. The remainder is immediately transported into the duodenum via the common bile duct. Stimulation of the muscular wall of the gall bladder to contract (by cholecystokinin released from the duodenal mucosa) produces a gradual release of bile that lasts for several hours. The bile then enters the duodenum by rhythmic relaxations of the sphincter of Oddi. Emptying of the gall bladder is very variable and almost never complete.3 An empty gall bladder during ultrasonography is an abnormal finding. Opioid drugs can cause complete closure of the sphincter of Oddi, so that during anesthesia the gall bladder may appear dilated; this usually resolves after anesthesia. Dilation of the larger bile ducts and /or the common bile duct indicates abnormalities such as cholangitis or bile duct obstruction.
7.1.2 Bloodsupply
The blood supply of the liver is both arterial and venous. Arterial blood comes from the hepatic artery, while venous blood is supplied through the portal vein. The outflowing blood is collected by a number of hepatic veins that enter into the inferior vena cava near the diaphragm. The total blood flow to the liver accounts for about 20-25% of the cardiac output (100-130 ml/min/100 g liver tissue). The portal vein accounts for 70% of the total blood flow to the liver and separates into two main branches entering the liver at the hilus. The right branch of the portal vein supplies the right side of the liver, while the bigger and longer left branch supplies the left and central portions of the liver. The intrahepatic portal vein divides into smaller branches, with the smallest branches ending in the portal areas.
Histologically, the terminal branches of the portal vein are the largest structures of the portal areas. These terminal branches end in inlet venules, which penetrate the periportal cell layer, the so-called limiting plate, where itdistributes the portal blood to the sinusoids. The hepatic artery also enters the liver at the hepatic hilus. Its branches accompany the portal vein. Terminal arterial branches end into the periportal sinusoids and connect with the venous and sinusoidal system. The biliary system of the liver is only supplied with arterial blood, while the parenchyma receives both portal and arterial blood. The sinusoidal blood enters the central veins, also known as terminal hepatic veins, in the liver lobule. The central veins then drain into branches of the hepatic vein. Finally, several branches of the hepatic vein enter into the caudal vena cava.4
The ratio between portal and arterial blood supply is not constant but varies with both physiological and pathological conditions. Portal blood flow increases following food ingestion, whereas the arterial blood supply increases when the portal blood flow is insufficient and decreases in the case of hepatic venous congestion. In patients with diseases that are associated with a severely decreased or total lack of portal blood flow (i.e., congenital portosystemic shunts, portal vein thrombosis), the arterial blood flow may increase by 100%, but even then the total hepatic blood flow remains subnormal. However, without this compensatory arterial blood flow the liver would not be able to survive. Histologically, the arterialization of the liver becomes visible after about 2 weeks. The hepatic arteries respond to the hypoperfusion of the liver and increase their blood flow. In addition, the arterial branches become tortuous and hypertrophic. Histologically, this can be recognized as an increased number of arterial sections in the portal areas. In the parenchyma, portal hypoperfusion causes hepatocellular atrophy.
In comparison, changes in arterial blood flow do not affect portal blood flow.4 During periods of fasting, the hepatic artery and the portal vein each supply approximately 50% of the hepatic oxygen requirement. In the case of hypoxia, which can occur in patients with anemia, the hepatic blood supply remains unchanged. When the oxygenation of the liver becomes deficient, either due to reduced erythrocyte oxygen binding or due to decreased perfusion, the liver adapts by an increased efficiency of oxygen uptake from approximately 40% up to nearly 100%. Hypoxia does not affect the total perfusion of the liver or the ratio between arterial and portal blood supply. Due to the large volume and the compliance of the hepatic vascular bed approximately 10-15% of the total blood volume is contained in the liver. This may increase by 100% in patients with severe congestive heart disease. Also, the large hepatic blood reservoir plays a role in compensating for acute blood loss.
It is important to note that, at the histological level, the liver consists of functional hemodynamic units or acini that are different from the lobules that are mere anatomically descriptive units. The acinar concept, which was proposed by Rappaport, places the portal vein at the center of the acinus with several central veins, also known as terminal hepatic veins (THV), at the periphery. The arteries and portal vein branches enter the liver via the portal areas. Portal blood is distributed from the larger portal veins throughout the acinus and drains into the sinusoids via short perpendicular inlet venules, which have a sphincter function and regulate the portal blood flow into the capillary bed. The THV collect blood at the outlet side of the sinusoids. Small tributaries of the hepatic artery enter the sinusoidal system at different places. Their blood flow is also controlled by a sphincter, which intermittently allows the flow of arterial blood, thus regulating the difference in pressure between the arterial system (100-110 mmHg) and the portal (0-5 mmHg) and sinusoidal (5-10 mmHg) systems.
According to the acinar concept, the liver circulatory units are divided into three zones, with the closest (Zone 1) around the portal area receiving blood with the highest contents of oxygen, growth factors, and nutrients, and Zone 3, which is farthest away from the portal area, and thus receives blood with the lowest concentrations of these compounds. In the case of sudden hypoxia or shock, when there is not sufficient time for compensation by increasing the oxygen binding of red blood cells or a more efficient oxygen uptake by the liver, the hepatocytes located in Zone 3 become necrotic. This results in centrolobular (Zone 3) hepatocellular necrosis. Such conditions are usually associated with increases in serum hepatic enzyme activities.
7.1.3 Microanatomy
The predominant cell type of the liver is the epithelial cell or hepatocyte. Hepatocytes make up 60% of the all the cells in the liver and since they are also the largest cell type found in the liver, they make up 80% of the liver volume (approximately 200,000 cells per mg hepatic tissue). Altogether, the liver contains about 100 billion hepatocytes. Other cell types that are present in the liver are endothelial cells, bile duct epithelial cells, Kupffer cells, and fat-storing cells. The latter cell type is also known as stellate cells.5
The hepatocytes are arranged in cell plates that are only one layer thick, radiating around the THVs. The free outer membranes on both sides of the hepatocyte are in contact with the sinusoids. Due to the numerous microvilli of that part of the membrane, the sinusoidal membrane occupies 70% of the overall hepatic cell surface area. A portion of the lateral membranes of adjacent cells form the bile canaliculi that are surrounded by tight junctions, which connect the hepatocytes on either side of the canaliculi. The canalicular membrane comprises approximately 15% of the hepatocyte membrane, and has a specialized excretory function. The sinusoidal membrane is separated from the sinusoids by one cell layer of sinusoidal
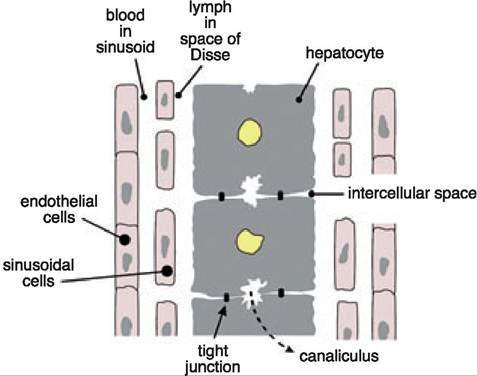
Figure 7.2:
Arrangement of cell types in the liver. This diagram shows the arrangement of endothelial cells, sinusoidal cells, and hepatocytes, which respectively form the conduits for blood, lymph, and bile flow. Hepatocellular necrosis leads to a direct connection between the canaliculi (i.e., the space of bile flow) and the space of Disse, resulting in a leakage of bile into the circulation (intrahepatic cholestasis).
Table 7.1: Hepatocyte functions in the different sublobular regions
Note that both lipid and protein metabolism are functions that are not localized to one region.
Zone 1: periportal area Zone 3: Centrolobular area The perisinusoidal space represents the beginning of the hepatic lymphatic system and leads lymph in a direction opposite to that of the blood flow, but in conjunction with the bile flow. The sinusoidal cells (endothelial and Kupffer cells) are very efficient in removing endotoxins and particles, such as bacteria. This, together with the many detoxifying metabolic functions of the hepatocytes and the biliary excretion of toxic substances, make the liver an important guardian against toxic substances entering the body from the GI tract. The bile in the canaliculi flows opposite to the blood flow into short collecting ducts (canals of Hering), which drain bile from the acini into bile ductules of the portal area. The luminal surface of these ductuli and the larger bile ducts are covered with cuboi- dal biliary epithelium.
Although hepatocytes are generally uniform and thus capable of performing most metabolic functions, there is some heterogeneity with respect to some functions. For example, enzymes that are part of the urea cycle are predominantly found in the hepatocytes of Zone 1 around the portal area. Also, the incorporation of ammonia in glutamine, which is regulated by glutamine synthetase, occurs exclusively in the hepatocytes around the central vein (THV). Carbohydrate metabolism also shows some degree of zonal organization, although it is not as strict as that of ammonia metabolism. Lipid and protein metabolism are not restricted to one particular zone. Drug metabolism via the cytochrome p450 mixed function oxidase system appears to be primarily located around the central veins. Copper accumulation in patients with copper storage disease mostly affects cells in Zone 3 in all dog breeds (Table 7.1).
Bilirubin excretion
Bile acid excretion, bile-salt-dependent bile production
Gluconeogenesis, glycogenolysis → glucose production
Ammonia detoxification through urea cycle enzymes
Glutathione production → protection against oxidative stress
Copper excretion
Bile-salt-independent bile production
Glycolysis, glyconeogenesis → glucose clearance
Ammonia detoxification through glutamine formation
Biotransformation; cytochrome p450 enzyme-dependant detoxification; conjugation reactions
cells (about 7% of the liver cells). Between the hepatocytes and the sinusoidal cell layers is the perisinusoidal space of Disse (Figure 7.2). The endothelial cell lining is fenestrated, and these openings permit even large molecules to be exchanged between the sinusoidal blood and the space of Disse. However, cells cannot pass through the fenestrations and remain in the circulation.
7.2