Bacterial Diseases
Enterotoxemia
Enterotoxemia, caused by C. perfringens, is recognized worldwide as a common, frequently fatal disease of goats. The disease also occurs in sheep and cattle, but species differences occur in the epidemiology, pathogenesis, clinical presentation, and management of enterotoxemia.
Distinctive characteristics of enterotoxemia in goats include the clinical prominence of diarrhea, the presence of severe enterocolitis at necropsy, and the frequent failure of vaccination to protect from clinical disease. The disease in goats has been reviewed (Uzal and Songer 2008; Sumithra et al. 2013).Etiology
Clostridium perfringens causes enteric disease in a wide range of species, including domesticated mammals, poultry, and humans. There are five main types of C. perfringens based on the lethal exotoxins that they produce, as presented in Table 10.5. While the main exotoxins traditionally recognized for typing the organism are alpha, beta, epsilon, and iota, it is now known that C. perfringens can produce up to 15 lethal toxins, including enterotoxin, beta-2 toxin, and perfringolysin O toxin, which can play a role in the pathogenesis of enterotoxemias in different livestock species. This discussion focuses on the disease in goats. Broader discussions of C. perfringens and the enteric diseases it produces in other species are available elsewhere (Songer 1996; Constable et al. 2017).
The principal cause of caprine enterotoxemia worldwide is C. perfringens type D. This Gram-positive, non-motile, spore-forming, toxin-producing, anaerobic, rod-shaped bacterium is resident in the ruminant digestive tract, generally in low numbers. It is passed in feces and may persist in soil, though it dies out more quickly in soil than other Clostridium spp. The organism has a quick generation time, as short as eight minutes, allowing for rapid proliferation in the intestine under favorable conditions.
It is the rapid proliferation of this resident organism and the associated release of toxins that cause disease in the host.C. perfringens type D produces two main toxins: alpha and epsilon. While both toxins are important for typing the organism, epsilon toxin is the principal virulence factor, as discussed later under pathogenesis.
C. perfringens type C has also been cited as a cause of caprine enterotoxemia in the United States and Britain, though definitive confirmation of its role is scarce (Barron 1942; Guss 1977). There is also a report of type C enterotoxemia in goats and sheep in Greece (Tarlatzis et al. 1963). The major toxins of type C are alpha and beta. Because beta toxin is degraded by trypsin, type C enterotoxemia is most common in very young animals with low concentrations of intestinal trypsin, usually under 10 days
Table 10.5 Clostridium perfringens types and the toxins produced.
Toxins produced (gene expressing the toxin)

a Expression of enterotoxin is not consistent within types. It is most often associated with type A isolates, but may be associated with other C. perfringens types as well.
b There is one reported case of caprine enterotoxemia involving a C. perfringens expressing alpha toxin and beta-2 toxin (Dray 2004). Similar findings were reported from cases of enterocolitis in horses (Herholz et al. 1999).
c The original identification of beta-2 toxin involved a C. perfringens type C associated with necrotic enterocolitis in a pig. The organism expressed alpha, beta, and beta-2 toxins.
of age. There is a single confirmed report of C. perfringens type E in goats, involving a 2-day-old kid with fatal diarrhea (Kim et al. 2013).
Enterotoxemia due to C. perfringens type B, which causes lamb dysentery, is also uncommon in goats. Type B contains alpha, beta, and epsilon toxins. In Iran, anomalous C.
perfringens type B strains (lacking hyaluronidase) were isolated from goats in two different but nearby outbreaks of clinical enterotoxemia (Brooks and Entessar 1957). In Germany, type B enterotoxemia was confirmed in a dairy goat herd and a herd of dwarf goats. Mortality of kids up to 12 days of age was reported following bouts of watery, yellow, bloodstained diarrhea (Scharfe and Elze 1995).In Greece, C. perfringens type A was cited as the most common cause of enterotoxemia in sheep and goats (Deligaris 1978). However, ascribing pathogenicity in goats to type A remains controversial, because it is considered to be the C. perfringens type most widespread in the intestines of warm-blooded animals and the environment and can rapidly overgrow in organs following death (Songer 1996). Another early report of type A enterotoxemia involved captive Siberian ibex (Capra sibirica) at a zoo (Russell 1970). Experimental challenge of domestic goats by intraduodenal administration of type A in broth culture produced only a transient diarrhea at 12 hours post inoculation, which resolved 12 hours later with no other clinical signs or deaths. In contrast, a second group of goats given type D in the same manner all developed diarrhea and convulsions and died within 36 hours (Phukan et al. 1997).
A fatal case of enterotoxemia in a 5-week-old Boer goat kid in Canada has been reported as being due to type A (Dray 2004). This was an unusual case in that the C. per- fringens isolated from the gut at necropsy was found to have genes that expressed alpha toxin and beta-2 toxin, making it an atypical type A isolate. Pathogenicity of type A is usually associated with alpha toxin alone. Beta-2 toxin has been characterized only more recently, originally in association with a C. perfringens type C organism (Gilbert et al. 1997). C. perfringens organisms that produce beta-2 toxin have now been associated with necrotic enterocolitis in piglets (Waters et al. 2003) and horses (Herholz et al.
1999), as well as this single reported case of caprine enterotoxemia (Dray 2004).Clostridium perfringens can also produce an enterotoxin (Songer 1996). Though most commonly associated with C. perfringens type A, it has been found in some isolates of types B and D as well. The gene for enterotoxin expression has been identified in type E isolates, but the toxin is not produced (Billington et al. 1998). The role of enterotoxin in the pathogenesis of enterotoxemia in goats has not been studied and its importance, if any, remains unknown.
Epidemiology
Enterotoxemia is frequently cited by producers, veterinarians, and extension personnel around the world as a common and important disease of goats. Paradoxically, there is little documentation of caprine enterotoxemia in the veterinary literature and comparatively little research that relates specifically to the disease in goats.
Descriptions of caprine enterotoxemia have come from Australia (Oxer 1956), Britain (Shanks 1949), Canada (Blackwell and Butler 1992), France (Delahaye 1975), South Africa (van Tonder 1975), Sri Lanka (Wanasinghe 1973; Ubeyratne and Jayaweera 2013), Switzerland (von Rotz et al. 1984), and the United States (Boughton and Hardy 1941; Guss 1977). Most outbreaks of caprine enterotoxemia involve dairy goats raised under intensive or semi- intensive management conditions. When fiber goats have been affected, as in South African or Texas Angoras, the disease is far more likely to occur during periods of corralling or restricted grazing in grain fields than during normal free-range grazing and browsing (van Tonder 1975; Ross 1981). The greatest losses caused by enterotoxemia in sheep occur in growing lambs on concentrate rations in feedlots, but this management situation is rarely encountered in goat practice.
Sudden changes in feedstuffs or feeding practices have been associated with triggering outbreaks of enterotoxemia in all affected species. Specific situations that make goats predisposed to this disease have included turnout to lush pasture, feeding of bread or other bakery goods, feeding of a bran/molasses mash to recently fresh does, excessive grain consumption after accidental access to feed storage sheds, and feeding of garden greens to goats unaccustomed to green feed (King 1980a).
Feed changes, however, are not a prerequisite for enterotoxemia to occur. The author (DMS) has observed an explosive outbreak of enterotoxemia in a herd of commercial antibody-producing goats that were fed an unchanging diet of hay and concentrate for many months before the outbreak.Abrupt changes in weather have also been associated with onset of enterotoxemia in other species, but this has not been well documented in goats. Nor has a strong seasonal pattern been identified. Intestinal tapeworm infections are believed to predispose feedlot lambs to enterotoxemia by slowing transit time of grain rations through the gut, allowing for more extensive proliferation of clostridia. An association between tapeworm infestation and predisposition to enterotoxemia in pastured goats has also been suggested, but additional studies are required (Rao 2011).
While sporadic cases of caprine enterotoxemia are common, outbreaks of enterotoxemia in goat herds with high morbidity rates also occur. On some premises, the outbreak of enterotoxemia can become well established, with new cases occurring over several weeks or months. The epidemiologic factors contributing to this disease pattern are unknown. A buildup of C. perfringens type D in the environment from earlier diarrheic cases is probably contributory.
Pathogenesis
Specific studies on the pathogenesis of enterotoxemia in goats are limited. In other ruminant species it is believed that commensal C. perfringens type D organisms reside in the gut without producing much damage, because bacterial numbers are low and any toxins that are produced are moved quickly through the gut by normal peristalsis. Sudden ingestion of readily fermentable, carbohydrate- rich feeds permits more undigested starch to pass through the rumen to the abomasum and intestine, where it serves as a nutrient substrate for rapid proliferation of the organism. Excess carbohydrate intake may also predispose to reduced motility. This proliferation of C.
perfringens type D in conjunction with reduced peristalsis enhances the concentration and pathogenic potential of the epsilon toxin produced by the organism. Epsilon toxin, after being converted from the prototoxin by intestinal trypsin, increases vascular permeability in the gut, thereby facilitating its own absorption into the bloodstream. A generalized toxemia ensues. The toxin is necrotizing, and specifically neurotoxic. Death is attributable to damage of vital neurons, generalized toxemia, and shock (Kimberling 1988).There is evidence that the pathogenesis of enterotoxemia in goats is different in terms of the intestinal effects. In field cases, diarrhea is a predominant clinical finding in goats relative to other species, and at necropsy, marked enterocolitis is often present (Blackwell and Butler 1992). This enterocolitis is an infrequent finding in lambs and calves.
The predilection of the gut as a target organ in caprine enterotoxemia has been demonstrated experimentally. Lambs and kids given broth infusions of C. perfringens type D intraduodenally via a cannula showed markedly different clinical and pathologic responses (Blackwell et al. 1991). In general, lambs showed lethargy, overt neurologic signs, minimal diarrhea, and death. Kids showed more prominent diarrhea and abdominal discomfort, with fewer neurologic signs preceding death. At necropsy, intestinal lesions in lambs were limited to a moderate edema of the colon and watery gut content, while in kids a severe necrotizing colitis was seen grossly and confirmed histologically.
More recent experiments involving introduction of epsilon toxin into ligated gut loops of lambs and kids has suggested a possible mechanism for the different clinical responses in sheep and goats. At two hours post inoculation, kids had accumulated much larger volumes of fluid and sodium in the small intestinal lumen than lambs. It was hypothesized that the more rapid endotoxin-induced accumulation of fluid in the small intestine of goats could more quickly flush away bacteria and toxin, moderating the intestinal toxin absorption. In contrast, the delayed physiologic response in the small intestine in sheep might result in greater absorption of epsilon toxin from the small intestine of this species, leading to the neurologic signs, brain lesions, and lung edema that is more commonly observed in naturally occurring cases of enterotoxemia in sheep compared to goats (Fernandez Miyakawa and Uzal 2003).
Clinical Findings
Three distinct clinical forms of enterotoxemia are recognized in goats, namely peracute, acute, and chronic. The peracute form occurs more frequently in young goats than in adults. The clinical course is usually less than 24 hours and may go unnoticed. Finding one or more dead animals is often the first indication of peracute enterotoxemia in a herd. In milk-fed kids, it is frequently the larger, more robust, aggressively feeding individuals that are affected. In weaned kids, the history may reflect some recent change in feeding practices or an opportunity to overeat. Clinical signs include a sudden loss of appetite, profound depression, marked abdominal discomfort manifested by arching of the back and kicking at the belly, loud and painful screaming, and profuse watery diarrhea containing blood and shreds of mucus. Fevers of 40.5 °C (105 °F) are recorded. Affected goats quickly become weak and recumbent. They may show paddling or convulsions, but frequently they just lapse into a coma without excitatory signs. Death ensues within hours. Recoveries are rare, even with treatment. When the peracute form occurs in milking goats, the onset of signs may be foreshadowed by a sudden drop in milk production.
In the acute form, similar clinical signs are seen, but with less severity. Abdominal pain and screaming may be absent or reduced. Feces may first turn pasty or soft, but then become watery. The clinical course lasts from three to four days. Severe dehydration and acidosis, caused by the profuse diarrhea, become complicating factors in these cases. Spontaneous recoveries may occur, but most animals die if not treated. The acute form more often affects mature goats. It can occur in herds with a solid history of vaccination against C. perfringens type D, so enterotoxemia should not be ruled out on the basis of previous vaccination. A history of recent feed changes may or may not be elicited.
In the chronic form, intermittent, recurring bouts of illness are observed over several weeks. Mature animals are usually affected. These goats are dull and listless, with reduced appetite and milk production if lactating. There is progressive weight loss with intermittent episodes of pasty or loose feces. It has been emphasized that the chronic form is extremely difficult to recognize as enterotoxemia unless there is prior knowledge of acute or peracute cases in the herd (Shanks 1949).
Clinical Pathology and Necropsy
Reports on hematologic and clinical chemistry changes during caprine enterotoxemia are limited. The author (DMS) has recorded complete blood counts on goats with the acute form of the disease. There is a distinct trend toward neutrophilic leukocytosis. In 39 affected goats, the mean white blood cell count was 16 200/mm3, with individual counts as high as 47 700/mm3 recorded. Packed cell volumes (PCVs) as high as 57% were noted in cases with severe dehydration. Hyperglycemia, azotemia, and increased serum osmolarity may occur in the later stages of enterotoxemia (Blackwell et al. 1991), but they are not specific to this disease.
Reports on necropsy findings in caprine enterotoxemia also are limited. In a series of 10 goat necropsies, enterocolitis was the most consistent finding in enterotoxemia (Blackwell and Butler 1992). The gut lesions were hemorrhagic, fibrinous, or necrotic in character. Enterocolitis was followed in frequency by pulmonary edema, renal tubular necrosis, and edema of mesenteric lymph nodes. Hydropericardium, a common finding in sheep, was seen in only one goat. Glucose may be detected in bladder urine of affected goats, but is not diagnostic. If necropsy is performed shortly after death, the detection of soft or pulpy kidneys supports the diagnosis of enterotoxemia. However, if necropsy is delayed, the pulpy kidney loses its diagnostic significance, because kidney autolysis eventually occurs post mortem regardless of the cause of death. Gross lesions may be entirely absent in some cases of enterotoxemia (Blackwell 1983).
Cerebral microangiopathy is a prevalent finding at necropsy in sheep with the disease, and is considered pathognomonic in that species, but is less commonly identified in goats. This may reflect a failure to examine the brains of affected goats histologically, since neurologic signs are not prominent clinically in caprine enterotoxemia. When the lesions are actively sought in histologic examination of brain tissue, they may be found with some frequency (Uzal et al. 1997). In one review of 44 cases of caprine enterotoxemia post mortem, only 1 goat had gross lesions in the brain, but 8 (18%) showed microscopic brain lesions, characterized by intramural vascular proteinaceous edema (Ortega et al. 2019).
In experimentally induced type D enterotoxemia in goats, produced by administration of starch into the abomasum along with duodenal instillation of either whole cultures, culture supernatant, or washed cells, goats developed diarrhea and a necrotizing colitis, as is typical of the species. They also developed lesions similar to those found more often in ovine enterotoxemia, including lung edema and cerebral vasogenic edema (Uzal and Kelly 1998).
Abnormal-appearing areas of bowel mucosa can be used to support the diagnosis of enterotoxemia. Impression smears of affected mucosa reveal a predominant population of large Gram-positive rods typical of C. perfringens (Figure 10.5). A similar preponderance of large Grampositive rods plus spores may be seen in smears made from diarrheic feces ante mortem.
Swabs from intestinal lesions can be submitted for anaerobic culture. Because C. perfringens type D may be isolated from the gut of normal goats, culturing the organism from the gut has been considered by some to have little diagnostic significance. However, survey studies of abomasal and intestinal culture from normal, healthy goats indicate that only 61% of goats sampled carried any type of enteric clostridial organism, and only 3% carried C. perfringens type D (Sinha 1970). Therefore, the isolation of the organism from swabs of necrotic intestine in animals dying of signs typical of enterotoxemia may have more diagnostic significance than is usually ascribed to it.
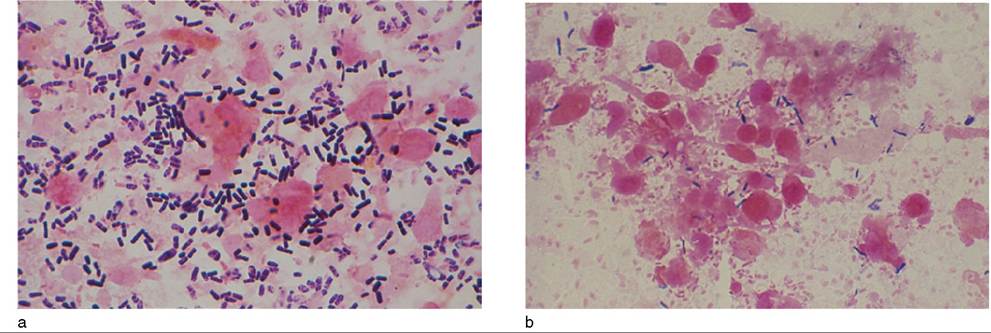
Figure 10.5 (a) Gram-stained impression smear of intestinal mucosa of goat with enterotoxemia. Note the abundance of large Gram-positive rods compared to (b) an intestinal smear taken from a normal goat at necropsy. Source: Courtesy of Dr. David M. Sherman.
The most convincing indication of enterotoxemia, however, is the presence of epsilon toxin in diarrheic feces or intestinal content. At least 10 mL of intestinal content should be submitted, taken from the areas of the bowel exhibiting lesions. The toxin is fragile, so gut content should be sent to the lab refrigerated or frozen. When type D enterotoxemia is suspected, intestinal content should be collected as soon as possible after death and frozen for shipment, because the epsilon toxin is especially labile.
Traditionally, toxin types have been identified by in vivo toxin-neutralization assays using mouse lethality and guinea pig or rabbit dermonecrosis as endpoints. However, alternative techniques are now available and in use for detection of the epsilon toxin associated with type D enterotoxemia. Among these are ELISA, CIEP, and latex agglutination tests. While ELISA tests have been found to be most sensitive, confirmation of the presence of epsilon toxin in small intestinal content by any of these methods can be considered as indicative of type D enterotoxemia, though a final diagnosis should be based not only on toxin testing but also clinical signs and postmortem findings (Uzal et al. 2003; Uzal and Songer 2008).
Beginning in the 1990s, PCR tests have been developed that identify the presence of the bacterial genes responsible for the production of the various C. perfringens toxins, thus obviating the need for direct identification of toxin for typing the organism (Daube et al. 1994; Meer and Songer 1997). Multiplex PCR genotyping is now routinely available in some diagnostic laboratories and can be applied to identify and type C. perfringens strains present in the gastrointestinal content or feces of goats (Uzal et al. 1996) or from paraffin-embedded tissues of goats (Warren et al. 1999). However, it should be noted that confirming the presence of the gene responsible for production of epsilon toxin does not confirm that epsilon toxin was responsible for producing disease in individual cases. The characteristics of epsilon toxin have been reviewed (Stiles et al. 2013).
Diagnosis
Diagnosis is based on a combination of characteristic clinical history and signs, evidence of enterocolitis at necropsy, isolation of C. perfringens from the feces or gut lumen, and identification of epsilon toxin or the gene expressing epsilon toxin in C. perfringens cultures from feces or gut lumen content. It has been suggested that a presumptive antemortem diagnosis of enterotoxemia can be made by observation of clinical improvement in response to administration of C. perfringens type D-specific antitoxins (Guss 1977). However, this has been reported as unreliable by others (Blackwell and Butler 1992).
Differential diagnosis of the peracute form must include all causes of sudden death, with particular emphasis on plant and chemical toxicities. Peracute deaths in kids younger than 3 weeks of age are not likely to be due to type D enterotoxemia, because normal gut trypsin levels may be inadequate to activate the epsilon prototoxin. Type C enterotoxemia, however, can cause death in these young kids.
When diarrhea is present in peracute or acute disease, the differential diagnosis should include coccidiosis, salmonellosis, yersiniosis, and in younger animals cryptosporidiosis and colibacillosis.
In milking does, the early stages of the acute form of the disease may mimic milk fever or hypocalcemia, but favorable response to administration of parenteral calcium salts in cases of milk fever should clarify the diagnosis. The chronic form of enterotoxemia and chronic salmonellosis may be clinically undifferentiable and diagnosis depends on bacteriologic methods.
Treatment
The prognosis for recovery is guarded in caprine enterotoxemia, even with treatment. Aggressive intervention is required. Affected animals should be managed in a warm and dry hospital pen. Intravenous fluid therapy providing mixed electrolyte solutions with bicarbonate is indicated in peracute and acute cases to counter shock, dehydration, and acidosis. Non-steroidal anti-inflammatory drugs such as flunixin meglumine (1 mg/kg intravenously every 12 hours) may be helpful in stabilizing animals in toxemic shock and will alleviate pain.
Commercially available type C and D antitoxins should be administered parenterally, preferably intravenously, in severe cases. Though recommended prophylactic doses are usually around 5 mL, therapeutic doses up to 100 mL have been administered. Because the antitoxin products are relatively expensive, a minimum effective dose is desirable. In Australian practices 15-20 mL has been effective (King 1980a). In those cases responsive to antitoxin, a rapid though sometimes temporary improvement may be noted within one to two hours. Equal or decreased doses can be repeated every three to four hours until the animal's condition has obviously stabilized. In the chronic form of the disease, two doses of 20 mL of antitoxin given four days apart was reported to be a reasonably effective treatment (Quarmby 1946). Diarrhea ceased in affected does and their milk production and body condition improved.
Decades ago, there were reports of allergic protein sensitivities to antitoxin observed in Saanen goats after repeated administration (Quarmby 1947). Though this may reflect impurities in older serum products, the potential for anaphylaxis, particularly in the Saanen breed, should be anticipated, and epinephrine (0.03 mg/kg intravenously) kept close at hand.
Antibiotic therapy may be helpful in reducing bacterial proliferation. Oral sulfas have been used successfully. To ensure that the drugs reach the abomasum and intestine, pretreatment with 5 cc of copper sulfate solutions (1 tablespoon CuSO4/liter of water) to close the esophageal groove followed rapidly by antibiotic administration has been recommended (King 1980a). Parenteral antibiotics including penicillin, ciprofloxacin, tetracyclines, or trimethoprimsulfa combinations may also be useful. C. perfringens isolates are often resistant to aminoglycosides (Songer 1996).
To limit the uptake and promote elimination of intestinal epsilon toxin, various cathartics and adsorbents have been given orally, including activated charcoal, magnesium sulfate, magnesium hydroxide, caffeine, and kaolin/pectin. Though the rationale for use is valid, therapeutic efficacy has not been proven.
In the face of outbreaks of enterotoxemia, all animals on the premises should be considered at risk. Previously vaccinated goats should be boostered, and unvaccinated goats should receive a prophylactic dose of antitoxin in conjunction with an initial vaccination to be boostered two to three weeks later. Any excessive feeding of carbohydrate should be curtailed immediately.
Control
Goats are considered highly susceptible to enterotoxemia and it is universally recommended that all goats be vaccinated against the disease as part of any basic caprine herd health program. At the same time, it is recognized that vaccination does not afford the same level of protection to goats that occurs in sheep, and that the persistence of serum antibodies against toxin at protective levels in goats is limited (Shanks 1949; Blackwell et al. 1983).
This species difference has been experimentally demonstrated. When sheep and goats were given equivalent doses of three different multivalent clostridial vaccines that included C. perfringens type D, levels of serum anti-epsilon toxin antibodies increased significantly in sheep and remained significantly higher than prevaccination levels at 28 days after vaccination. In goats, only one of the same products produced a significant increase in serum antiepsilon toxin antibody titers, and titers returned to prevaccination levels for all products by 28 days (Green et al. 1987).
The required level of anti-epsilon toxin antibody in serum that protects goats from clinical enterotoxemia is not fully established, but recent research suggests that it is likely 0.25 IU∕mL (Uzal and Kelly 1998; Uzal et al. 1998). In one study, goats vaccinated with an incomplete Freund's adjuvanted vaccine had high anti-epsilon antibody titers between 2.45 and 230 IU∕mL. They showed no clinical signs or postmortem lesions after intraduodenal administration of C. perfringens type D culture supernatant. In contrast, goats receiving a commercially available aluminum hydroxide adjuvanted vaccine only produced antibody levels in the range of 0.22-1.52 IU∕mL, and four of five goats developed diarrhea and some evidence of colitis at necropsy, while unvaccinated control goats all developed severe enterocolitis (Uzal and Kelly 1998). It seems, therefore, that the occurrence of enteritis in vaccinated goats derives from the fact that conventional commercial vaccines, which are generally approved for sheep, may not produce sufficiently high antibody responses in goats to effect protection against the enterocolitic form of the disease common in goats (Uzal and Kelly 1998).
In a second study, antibody levels were measured after vaccination with a standard commercial vaccine following a single injection alone, or after booster injections given either 28 days or 42 days later. In all three protocols, antibody levels had dropped below the putative protective threshold of 0.25 IU/mL in most of the goats by 98 days after the initial vaccination (Uzal et al. 1998).
These findings support the recommendation that goats should be vaccinated at intervals of three to four months to maintain adequate protection against enterotoxemia using currently available vaccines, especially in herds where there is a history of enterotoxemia. At the least, goats should be vaccinated semi-annually. Initial vaccinations should be followed by a booster vaccination three to six weeks later and semi- or tri-annual vaccinations should be timed so that the last vaccination occurs two to three weeks before parturition in pregnant does. This enhances the protective effect of colostrum for suckling kids. Kids should be vaccinated initially at four to six weeks of age and certainly before weaning. They should also be boost- ered three to six weeks later, depending on product recommendation.
Vaccines limited to C. perfringens types C and D with or without tetanus may be preferable to the use of more complex, polyvalent clostridial vaccines commonly approved for sheep and cattle. These polyvalent vaccines may be more expensive and provide unneeded protection against clostridial diseases that occur uncommonly in goats. Furthermore, in at least one study, anti-epsilon toxin antibody titers were sometimes significantly lower in goats in response to a multivalent vaccine than to a bivalent C. perfringens types C and D vaccine (Blackwell et al. 1983). Few products are approved specifically for goats, and in general sheep dosages are used.
There is a marked tendency for goats to develop local tissue reactions at the vaccine injection site. This is quite troublesome in show goats, and also complicates caseous lymphadenitis control, because owners may have trouble differentiating between lymph node abscesses and injection site reactions. The occurrence of sterile abscesses of 2-5 cm in diameter after clostridial vaccination is well documented and does not necessarily result from careless injection technique, but rather is intrinsic to some vaccine preparations (Smith and Klose 1980; Blackwell et al. 1983; Green et al. 1987).
Scrupulous vaccination technique should be observed when vaccinating show goats to avoid any implication that technique was at fault, should abscesses develop. Use a new needle for each vaccination. Subcutaneous vaccination in the loose skin on the chest wall behind the elbow is recommended as a site where reactions will be less visible in show goats. However, if reactions occur at this site and become secondarily infected, they may grow quite large before they are noticed. In commercial goats, therefore, vaccination on the neck is recommended. Practitioners should sample several vaccine products to determine which produces the least tissue reaction. No correlation between tissue reaction and immunogenicity has been demonstrated (Green et al. 1987).
In addition to vaccination, control of enterotoxemia depends on avoidance of sudden feed changes, vigilance against overeating, and guarding against accidental access to grains and other stored feeds. Tapeworm control may also be beneficial.
SaLmoneLlosis
Salmonellosis continues to be an important infectious disease for several reasons: changing patterns of infection and morbidity rates in virtually all domestic species of livestock and poultry, public health concerns regarding human illness from animal sources, increased occurrence of multiple drug resistance, and economic losses to producers and food processors caused by condemnations and adverse publicity relating to the zoonotic potential of the infection. These issues are all relevant to the goat industry.
Etiology
Salmonella are Gram-negative, non-spore-forming, rodshaped bacteria in the family Enterobacteriaceae. There are more than 2400 serotypes, also known as serovars. On a worldwide basis, the serovar S. Typhimurium is the most common cause of the diarrheal and septicemic forms of clinical salmonellosis in cattle, sheep, horses, and swine. This serovar has been associated with clinical salmonellosis in goats in the United States (Bulgin and Anderson 1981). In two British reports, however, S. Dublin was isolated from sick goats (Levi 1949; Gibson 1957), while in a Nigerian investigation of diarrheic kids, S. Poona was the sole isolate identified (Falade 1976). In an Australian outbreak, four serovars were isolated from goats dying with severe diarrhea: S. Adelaide, S. Typhimurium, S. Muenchen, and S. Singapore (McOrist and Miller 1981). Isolates from cases of gastroenteritis and septicemia in goats in India include S. Typhimurium, S. Bere, S. Colombo, S. Newport, S. Tennessee, and S. Worthington (Janakiraman and Rajendran 1973). S. Enteriditis, S. Abony, and S. Cerro were cultured from cases of kid diarrhea in Greece (Zdragas et al. 2000).
Salmonella can also cause caprine abortion. Abortions may occur as a sequela to septicemia with any Salmonella serovar, but a specific epizootic abortion syndrome of goats and sheep caused by S. Abortusovis occurs in Mediterranean and Middle Eastern countries (Leondidis et al. 1984). S. Abortusovis may also cause enteric salmonellosis (Sanchis and Cornille 1980). Salmonella are an infrequent cause of mastitis in goats, gaining entry to the mammary gland through the teat canal from a contaminated environment.
Numerous Salmonella serovars have been isolated from the feces of healthy goats (Kapur et al. 1973; Abdel-Ghani et al. 1987) and more than 40 serovars from the viscera of healthy goats at slaughterhouses around the world (Nagaratnam and Ratnatunga 1971; Kumar et al. 1973; Gupta 1974; Arora 1978; Nabbut and Al-Nakhli 1982; Subasinghe and Ramakrishnaswamy 1983; Faraj et al. 1983; Diaz-Aparicio et al. 1987; Woldemariam et al. 2005; Chandra et al. 2006). S. Paratyphi, the cause of paratyphoid in humans, has been isolated from mesenteric lymph nodes of slaughtered goats in the Middle East and Asia, suggesting that the goat can act as a source of infection for humans. In slaughterhouse surveys of viscera, Salmonella are most often isolated from mesenteric lymph nodes and gall bladders.
Epidemiology
The occurrence and outcome of salmonellosis in livestock can be influenced by a number of factors. These include the existence of host-adapted Salmonella serovars, the development of a carrier state, impaired immunity of the host, the occurrence of severe stress including prolonged transport or deprivation of food and water, intensive management practices, and exposure to contaminated feeds or infected animals. The role of all these various factors in caprine salmonellosis is not fully described.
There are no known host-adapted Salmonella serovars in goats. S. Dublin, which is considered to be host adapted to cattle in Britain, caused clinical disease in goats, but there was no indication that a carrier state or persistent shedding occurred in exposed or recovered goats based on repeated fecal cultures or tissue cultures at necropsy (Levi 1949; Gibson 1957).
Reports of naturally occurring salmonellosis indicate that latent carrier states occur in goats, and that latent infections can progress to active fecal shedding of Salmonella or to overt clinical disease in response to stress of transport, excessive handling, feed or water deprivation, sudden feed changes, or parturition. The frequent isolation of Salmonella from the organs of healthy goats at slaughter, as cited above, also indicates the existence of a carrier state.
A latent carrier state in goats has been demonstrated experimentally with S. Typhimurium (Arora 1983). Goats challenged orally showed peak fecal shedding at three days after challenge, but all shedding ceased by two weeks. During the three subsequent weeks, no organisms were recovered from feces. However, when these goats were later stressed by transport, 60% resumed fecal shedding of S. Typhimurium. The organism was isolated from mesenteric lymph nodes, livers, and spleens at necropsy. In contrast, attempts to confirm the existence of a nasal carrier state in kids from premises with a history of salmonellosis were unsuccessful, though calves and piglets on the same premises yielded the organism (Garg and Sharma 1979).
A carrier state in goats has been demonstrated under field conditions in Australia, where 26% of fecal samples collected from feral goats yielded S. Typhimurium when the samples were taken immediately after capture and transport to a feedlot. In contrast, the frequency of positive samples was 4.8% one month later, 1.6% two months later, and 0% three months later, suggesting that the stress of capture and transport triggered fecal shedding of S. Typhimurium and the frequency of shedding declined as the goats became adapted to their surroundings (Al-Habsi et al. 2018).
Transmission is most commonly by the fecal-oral route. The introduction of unrecognized carrier animals into susceptible populations is of major importance in the spread of salmonellosis. Cross-species infection can also occur, as reported in the transmission of S. Dublin from calves to goats via fecal-contaminated pens and bedding, and of S. Typhimurium from ducks to goats via consumption of contaminated water (King 1980c). Other potential sources of infection for goats include raw sewage, contaminated feedstuffs, rodents, birds, and other livestock species, although documentation of such transmissions to the goat is rare.
Other factors that may predispose to clinical salmonellosis in goats include close contact with active shedders, excessive buildup of organisms in the environment caused by poor hygiene or poorly constructed feeders and waterers, concurrent disease, particularly severe gastrointestinal parasitism, stress of capture in feral goats, and, in neonates, failure of transfer of passive immunity from the does' colostrum. There is little documentation that intensive management with many goats maintained in confinement housing increases the risk or spread of salmonellosis, although this is considered a major factor in regard to dairy cattle.
Human infections with Salmonella from goat products have been documented. Raw or undercooked goat meat can pose the risk of salmonellosis, and poor hygiene at abattoirs can contribute to the contamination of goat carcasses. In an abattoir study at different sites in North America, Salmonella were isolated from fecal samples taken directly from the distal colon following evisceration at slaughter in 10.3% of the goats sampled (Hanlon et al. 2018). In an abattoir study at two sites in Australia, Salmonella were isolated from 46.3% of the fecal samples taken from goats in a similar fashion at slaughter (Duffy et al. 2009).
Goat milk or goat milk cheese, especially if unpasteurized, can also be a source of Salmonella infection for humans. Severe gastroenteritis and even fatalities can occur. In France in 1993, a commercial goat cheese made from unpasteurized goat milk led to 273 human cases of salmonellosis, with one death. Though the processing plant derived milk from 40 goat herds, the offending organism, S. Paratyphi B, was traced back to a single goat farm, and 33 tons of cheese stored at the plant had to be destroyed (Desenclos et al. 1996). In a separate incident, three brands of unpasteurized goat cheese derived from a single herd of 260 goats in France were associated with 52 cases of human disease in seven European countries: France, Switzerland, Sweden, Austria, Germany, the United Kingdom, and the Netherlands. The causative agent was a relatively rare sero- var, S. Stourbridge (Espie and Vaillant 2005). Milk samples from all goats in the source herd were cultured in the ensuing epidemiologic investigation and the organism was found in the milk of only one asymptomatic carrier. This outbreak underscores, among other things, how widely zoonotic disease can be disseminated under modern systems of marketing and trade. In France, 25 cases of human infection with the uncommon serotype S. Muenster occurred in association with the consumption of unpasteurized goat milk cheese from a single producer (van Cauteren et al. 2009). Signs of infection included fever, diarrhea, nausea, and abdominal pain.
Pathogenesis
The pathogenesis of enteric and septicemic salmonellosis has not been specifically studied in goats and is presumed to be similar to that described in other species (Constable et al. 2017). After ingestion, the organism becomes established in the intestines, most commonly in the ileum. A marked enteritis develops, most likely as a result of release of endotoxin from dying organisms. Diarrhea results at least in part from the inflammatory enteritis, but the organisms may also elaborate enterotoxins that have a hypersecretory effect on villous epithelial cells, leading to accelerated fluid and electrolyte loss from the gut.
Salmonella are invasive bacteria and can penetrate the mucosa, gaining entry to the lymphatics. Several outcomes are possible. In individuals with decreased disease resistance or severely impaired immunity, the organism can then gain entrance to the blood, producing a generalized septicemia with marked endotoxemia and usually death, while animals with a strong immune system may clear the infection entirely. In animals with moderate disease resistance, a transient bacteremia may occur with subsequent localization of the organism in liver, gall bladder, spleen, and mesenteric lymph nodes. These animals become latent carriers, and in the face of subsequent stresses they may develop clinical septicemia or enteritis, or they may remain subclinical but initiate fecal shedding of the organism, usually as a result of reinoculation of the gut via the infection in the gall bladder. In carrier does that are pregnant, the stress of parturition may induce bacteremia and result in exposure of the kid either in utero or via the milk, or by fecal contamination of the udder.
Animals experiencing infection may produce detectable antibodies against flagellar (H) and somatic (O) antigens of Salmonella. In goats experimentally infected with S. Typhimurium, antibodies to O and H antigens were detectable by five days post infection in one study (Sharma et al. 2001) and not until after seven days in another (Otesile et al. 1990). The flagellar antibodies appear to be more persistent in goats. The role of antibody in subsequent immunity is not clear. Immunity to the same strains of Salmonella may develop after initial exposures, but cross-immunity is limited. This fact has impeded the development of broadly efficacious vaccines.
Clinical Findings
Three age patterns of salmonellosis have been identified in goats, namely neonatal septicemia during the first week of life, preweaning enteritis in kids 2-8 weeks of age, and enteritis/septicemia of mature goats. The prognosis is grave in the first, poor in the second, and guarded in the third.
In neonatal septicemia, kids usually appear normal at birth and may die acutely by 36 hours, with no signs other than depression. Occasionally, signs of gaseous abdominal distension with pain or diarrhea may be seen.
In enteritis of older kids, there is an acute onset of depression and anorexia. A profuse, watery, foul-smelling, yellow to greenish-brown diarrhea develops, accompanied by fever up to 41.7 °C (107 °F). Affected kids rapidly become severely dehydrated, weak, and recumbent. Some may die within 8 hours of the onset of diarrhea, with most dying within 24-48 hours. Fever may subside after 24 hours and temperature can become subnormal as animals develop shock. Morbidity and mortality rates can be high, particularly as kid births are often concentrated into a short period and numerous kids on the premises may be at risk.
The adult form of the disease tends to be more sporadic, with lower morbidity and mortality rates. Affected goats become acutely depressed, anorexic, and febrile, and develop a watery, foul-smelling diarrhea that can be yellow, gray, or greenish-brown. Dehydration and weakness rapidly ensue and death can occur within 24-48 hours of the onset of signs. A chronic form of adult disease can also occur in which similar but milder signs are seen with subsequent recovery, followed by intermittent, recurring bouts of diarrhea. These animals tend to become progressively emaciated and may develop anemia.
In published cases of caprine salmonellosis, bloody diarrhea or fibrinous casts are rarely reported. This is in marked contrast to the disease as described in cattle or sheep.
Clinical Pathology and Necropsy
Goats with salmonellosis may have a marked leukopenia early in the course of disease followed by a rebound leukocytosis, with neutrophilia and a left shift in animals that do not die acutely. Severe metabolic acidosis can be expected, along with losses of sodium and potassium caused by diarrhea. Liver-specific enzymes may be elevated in septicemic salmonellosis due to foci of infection in the liver. In chronic salmonellosis, anemia and hypo - proteinemia may be observed. In neonates, serum immunoglobulin levels may be low if failure of passive transfer of antibody from the dam's colostrum has predisposed to clinical salmonellosis. Bacterial culture of the doe's milk and feces is also indicated to identify the dam as the pos - sible source of infection.
For a definitive antemortem diagnosis in enteritis cases, repeated attempts at fecal culture should be made. Appropriate enrichment media should be used. In some cases, attempts to culture the organism from diarrhea are unsuccessful, but positive culture results may be obtained from formed feces after the diarrhea has resolved. Subsequent biochemical characterization and serotyping of isolates at a reference laboratory are recommended because of the different zoonotic implications of various Salmonella serovars. Blood cultures may be attempted on septicemic cases before antibiotic therapy, and antibiotic susceptibility testing of isolates should be requested, because the occurrence of antibiotic resistance in Salmonella is increasing.
Serology may be of some value in the diagnosis of acute salmonellosis, particularly in evaluating a herd outbreak. In outbreaks of S. Dublin infection, goats had strong persistent antibody responses to flagellar H antigens of homologous S. Dublin for several months after exposure. Non-exposed control goats had background agglutination titers up to 1 : 80, whereas exposed goats had titers as high as 1 : 20 480.
At necropsy, kids dying acutely of septicemia may show minimal gross lesions consisting of some hemorrhage on serosal surfaces, increased pericardial and peritoneal fluid, and gas-distended bowel. Histologically, edema of the abomasum and dilation of the tips of the intestinal villi have been observed. Culture should be attempted from multiple organs.
In older kids and adults with enteritis, gross lesions can be more apparent and include petechial hemorrhage of serosal surfaces, pericardial and peritoneal effusions, enlarged and edematous mesenteric lymph nodes, congestion of the lungs and liver, edematous thickening of the gall bladder, and marked diffuse inflammation of the intestinal mucosa. The small and large intestines can be involved and the inflammation can range from catarrhal, to hemorrhagic, to necrotic, with diphtheritic membranes present in more prolonged cases. In chronic cases, lack of abdominal fat, miliary nodules in the liver, and hepatic lipidosis may also be seen.
Histologically, the intestinal inflammation is confirmed. The small intestine may show dilated mucosal crypts containing cellular debris and neutrophils, submucosal infiltrates of neutrophils and lymphocytes, and edema of the lamina propria. There may be small foci of hepatic necrosis containing aggregates of mononuclear cells. It is reported from experimental infection in goats with S. Typhimurium that histologic lesions in mesenteric lymph nodes may be severe, even when enteric lesions are not conspicuous (Sharma et al. 2001). In goats, the mesenteric lymph nodes are the most promising source for bacterial isolation. Intestinal content, spleen, liver, and bile may also be cultured successfully.
Diagnosis
In newborn kids, starvation, hypothermia, lethal congenital defects, and colisepticemia must be considered in the differential diagnosis. In older kids with acute enteritis, the major differential diagnoses are coccidiosis, cryptosporidiosis yersiniosis, enterotoxemia caused by C. perfringens type D, and, if pastured, gastrointestinal nematodiasis. In adult animals, enterotoxemia and gastrointestinal nemato- diasis should be ruled out. PPR must be considered in countries where it occurs.
Treatment
Treatment should be directed at rehydration and maintenance of circulatory volume, correction of acid-base and electrolyte imbalances, amelioration of endotoxic effects, and the control of bacteremia when present. Intensive fluid therapy is indicated, and it is unlikely that any route other than intravenous will be of value. Balanced electrolyte solutions supplemented with sodium bicarbonate and potassium are indicated. In septicemic neonates, glucose supplementation is also advisable. If neonates are hypogammaglobulinemic, whole blood or preferably plasma transfusions are desirable. Non-steroidal antiinflammatory drugs such as flunixin meglumine administered parentally can be helpful.
The use of antibiotics in the treatment of salmonellosis is controversial, because in other species it is associated with prolonged fecal excretion of bacteria, increased likelihood of developing a carrier state, and the promotion of antimicrobial resistance in the organism. These associations have had limited investigation in the goat. Two goat isolates made in India, S. Typhimurium and S. Weltevreden, were shown to possess R-factor that imparted multiple antimicrobial resistance to tetracycline, oxytetracycline, and chlortetracycline (Kumar and Misra 1983). An abattoir survey of 204 slaughtered goats in India yielded 60 Salmonella isolates with 40 patterns of antimicrobial resistance. All isolates were susceptible to chloramphenicol and imipenem, while 70% were resistant to nitrofurantoin and 52% to amikacin. Multiple resistance to three or more drugs occurred in 52% of the isolates, while only 8% of isolates were sensitive to all drugs tested; 13% were resistant to gentamicin and 18% to tetracycline (Chandra et al. 2006).
Given growing concerns about antimicrobial resistance, antibiotics should be used prudently in the treatment of salmonellosis, and whenever possible be based on microbial culture and antibiotic sensitivity testing. In the case of septicemic kids, it is unlikely they would have any chance of survival without the use of antibiotics. In addition, it is extremely difficult to establish clinically if there is a component of septicemia in advanced cases of enteritis where severe dehydration and electrolyte imbalance lead to weakness and recumbency, suggestive of endotoxic shock. The use of antibiotics in these cases is justified. The major concern is that antibiotic-treated animals should not be subsequently introduced into naive herds, because it is impossible to be sure that the carrier state has not developed.
Where its use is permitted, chloramphenicol, at a dose of 10 mg/kg intravenously every 12 hours, has proven to be a drug to which most caprine isolates of Salmonella are susceptible. Caprine isolates are also frequently susceptible to cephalothin, the aminoglycosides gentamicin and kana- mycin, and trimethoprim-sulfonamide combinations. The aminoglycosides must be used cautiously in these cases, because the severe dehydration that is often present can aggravate the inherent renal toxicity of these drugs.
Gentamicin is given intramuscularly or subcutaneously at a dose of 1 mg/kg every eight hours, and kanamycin at 5 mg/kg every eight hours. Trimethoprim is rendered inactive by rumen degradation and therefore trimethoprimsulfa combination drugs must be given intravenously or subcutaneously to adults. They should also be given intravenously to septicemic neonates to provide rapid plasma concentrations. They may be given orally to young preruminant kids with enteritis. The dose is 30 mg/kg orally once a day or 15 mg/kg intravenously every 12 hours. Sensitivities to tetracyclines, nitrofurantoin, sulfonamides, neomycin, streptomycin, erythromycin, and ampicillin are more variable. Resistance to penicillin is typical (Nabbut et al. 1981; Mago et al. 1982; Kumar and Misra 1983). In outbreaks, mass medication with sulfonamides or tetracyclines can be helpful if the isolate is susceptible to these drugs. Drugs should be added to the water rather than the feed, because many affected animals will not eat but may continue to drink.
Control
To prevent introduction of Salmonella into a naive herd, purchase of outside animals should be discouraged. If purchases are made, they should be directly from healthy source herds and not through sales barns or markets. Because the stress of transport and introduction to new surroundings may trigger fecal shedding or overt clinical disease, newly purchased animals should be isolated from the herd for at least three weeks after purchase and, if practical, bacterial cultures of feces should be performed, particularly if the animal develops diarrhea during isolation.
In the face of an outbreak, it is important to identify the source of infection. Because the source of Salmonella can be contaminated feed or water, subclinical carrier animals, newly introduced livestock, or other animals on the premises, including rodents and birds, extensive animal and environmental sampling may be necessary to identify and remove the offending source.
Affected animals and their cohorts should be isolated away from the rest of the herd and rigorous hygienic measures should be practiced when dealing with these animals. If affected animals have been moved from the original site of the outbreak, those premises should be thoroughly cleaned, repeatedly disinfected, and allowed to remain unused for several weeks. Common household bleach is an inexpensive and effective disinfectant, providing that organic material first has been removed from floors, pens, feeders, and waterers.
When sporadic cases or recurrent outbreaks of salmonellosis continue to occur in a herd, the presence of carrier animals must be considered. Repeated herdwide fecal culture testing with culling of shedders may be costly but effective. Strict attention must be paid to ensuring that kids receive adequate colostrum and that kidding facilities and feeding equipment are kept clean. Kids should be reared in a separate facility away from adults, and preferably in individual pens or hutches rather than in groups.
Vaccination can play an important role in the control of Salmonella infection in ruminant livestock, but at present the options are limited for goats, because there are no vaccines approved specifically for use in this species. A bivalent killed bacterin containing S. Typhimurium and S. Dublin is available for use in cattle in many countries, including the United States, but the effectiveness of whole cell killed Salmonella bacterins is limited. Vaccines based on the core lipopolysaccharide (LPS) common to Gram-negative bacterial cell walls are also commercially available for cattle and may offer better cross-protection between serovars, because the repeat polysaccharide units of the LPS that confer serotype specificity are not present in the vaccine.
There is active research on the development of new vaccine technologies to improve cellular and humoral responses and to overcome the general lack of crossprotection provided by monovalent vaccines against heterologous Salmonella serovars. One promising advance has been the development of a vaccine that produces antibodies against siderophore receptor sites and porin proteins found on the outer cell membrane of Enterobacteriaceae, including Salmonella. The receptor sites and proteins are necessary for the transfer of iron into the bacterial cell, a requisite for bacterial multiplication and survival. The receptor site structures are conserved across various Salmonella serovars, so cross-protection against multiple serovars by using this approach can be significant. A commercial vaccine based on this technology is now available in the United States, but is limited to use in cattle (Stevens and Thomson 2005). At present, the use of autogenous bac- terins made from Salmonella isolated on the premises can be considered for control of infection in known infected herds of goats. Even if vaccines do become available for use in goats, effective control programs for salmonellosis will not only depend on vaccination, but must also include hygienic management practices and the maintenance of closed herds.
Yersiniosis
Yersiniosis has emerged as a common cause of enteritis and death in goats in New Zealand, where it appears to have become enzootic. It is also sporadically associated with enteritis, abortion, mastitis, internal abscesses, and mortality caused by septicemia in goats throughout the world. Yersiniosis is a zoonotic disease.
Etiology
Yersinia spp. are Gram-negative, aerobic and facultatively anaerobic, non-lactose-fermenting coccobacilli of the family Enterobacteriaceae. They grow readily on blood agar and MacConkey's agar, but may be quickly overgrown, particularly when isolating from fecal specimens.
There are three pathogenic species in the genus Yersinia. Yersinia pestis causes plague in humans and rodents and will not be considered further, except to note that there are recorded cases of human plague traced to the slaughter, skinning, and butchering of sick goats. The goat is considered a sentinel animal in plague epidemics (Christie et al. 1980).
Yersinia enterocolitica and Yersinia pseudotuberculosis are pathogenic for humans and a range of animal species including goats. Y. enterocolitica is ubiquitous in the environment. Y. pseudotuberculosis is a common inhabitant of the gut of many domestic and wild animals. The conditions caused by either of these two species are known collectively as yersiniosis.
Different serotypes are recognized, based primarily on somatic O antigens. There are six main serotypes (I-VI) of Y. pseudotuberculosis. Serotypes I and especially III are most often associated with disease in goats (Hubbert 1972; Jones 1982; Hodges et al. 1984; Slee and Button 1990b; Seimiya et al. 2005). Serotype III produces an exotoxin that may be a virulence factor.
Y. enterocolitica isolates are classified according to biotype, which refers to their biochemical characteristics, and serotype, which refers to their antigenic characteristics, mainly associated with somatic O antigens. Because some antigens are common to both pathogenic and non- pathogenic isolates, ideally organisms should be described by first biotype and then serotype, though these full descriptions are not always included in the earlier literature. There are five main biotypes or biovars of Y. enteroco- litica, and many serotypes or serovars. Biotype 5 serotype 0:2,3 has been associated with caprine enteritis in Norway (Krogstad et al. 1972) and was also associated with enteric disease of goats and sheep in Australia and New Zealand (Slee and Button 1990a).
In standard serologic tests, such as the microagglutination test, Y. enterocolitica serotype 0:9 is cross-reactive with Brucella abortus and can confound surveillance in brucellosis control programs. It was demonstrated in Canada that normal goats may be infected with serotype O:9 and possess antibody that leads to false-positive brucellosis test results when using B. abortus antigen (Mittal and Tizzard 1980). An ELISA technique using refined antigens from B. abortus has been reported to serologically differentiate Brucella-infected animals from those with Y. enteroco- litica O:9 (Erdenebaatar et al. 2003).
Epidemiology
Many sporadic episodes of yersiniosis were reported in animals and humans throughout the world for most of the twentieth century. In recent years, gastrointestinal illness in humans caused by Y. enterocolitica is increasing, and links to animal contact and animal food sources are often suspected or confirmed (Nesbakken 2006). Y. enterocolitica has been isolated from raw goat milk offered for sale in Northern Ireland and Australia (Hughes and Jensen 1981; Walker and Gilmour 1986), and yersiniosis in a goat keeper was recorded in Norway during an epizootic of diarrhea in goats with Y. enterocolitica biotype 5 serotype O:2,3, possibly as a result of animal or fecal contact (Krogstad et al. 1972).
Nevertheless, goats are not considered to be significant carriers of human pathogens (Nesbakken 2006). A recent sampling survey of 575 goats in 24 goat herds in northern Germany yielded 17 Y. enterocolitica-positive fecal samples from five herds. All isolates were biovar 1A, which is considered to be non-pathogenic, and it was concluded that milk, cheese, or meat from these goats did not represent a risk of human yersiniosis (Arnold et al. 2006). However, a serosurvey in the same region of Germany during the same time frame indicated that sera from 66% of 681 goats from 28 farms had antibodies to Yersinia outer proteins (Yop) generally associated with pathogenicity (Nikolaou et al. 2005). One possible explanation is that goats are indeed often exposed to subclinical infections with pathogenic Yersinia, but mount effective immune responses that eliminate these infections. This explanation is supported by the findings of a longitudinal study in New Zealand wherein pathogenic Yersinia cultured from the feces of young goats could not be identified again on subsequent monthly samplings from those same goats (Lanada et al. 2005).
Epizootics in lab animal colonies and aviaries due to Y. pseudotuberculosis have been the most common form of the disease in animals, but livestock epizootics also occur (Obwolo 1976). Stress, overcrowding, and sudden cold weather can be predisposing factors. Birds and rodents are considered reservoirs of infection and may introduce virulent strains into susceptible herds by contaminating feedstuffs. Swine are a carrier source of Y. enterocolitica and may infect ruminant species when commingled. Because Yersinia organisms are often present in the gut of normal animals, factors that compro - mise gut mucosal integrity, such as concurrent parasitism or coarse feed-related abrasions or ulcerations, can predispose to septicemic yersiniosis.
Yersiniosis has been increasingly recognized as a cause of various disease problems in goats in the past two decades. Abortion and postparturient deaths in goats because of Y. pseudotuberculosis have been reported from Germany (Albert 1988), India (Sulochana and Sudharma 1985), Japan (Morita et al. 1973), and the United States (Witte et al. 1985). Liver abscesses and granuloma formation in goats because of Y. pseudotuberculosis have been documented in the United States, Japan, and Australia (Hubbert 1972; Morita et al. 1973; Slee and Button 1990b). Chronic mastitis in does caused by Y. pseudotuberculosis has been reported from California (Cappucci et al. 1978) and the United Kingdom (Jones 1982). Clinical mastitis caused by Y. enterocolitica has not been reported in goats, and efforts to induce it experimentally were unsuccessful (Adesiyun and Lombin 1989). A retrospective survey from veterinary diagnostic laboratories in California identified 42 cases of disease due to Y. pseudotuberculosis in goats over a 23-year period. This represented 0.9% of the 4455 goats necropsied during that time. In 27 cases, the diagnosis was enteritis or typhlocolitis, and all these cases occurred during colder and wetter periods of the year. Clinical presentations of yersiniosis in the remaining cases included abscesses in 6, abortion in 5, conjunctivitis in 2, and hepatitis in 2 (Giannitti et al. 2014).
Yersiniosis has become enzootic in goats in New Zealand. In a large study of goat mortality on the North Island, yersiniosis, mainly due to Y. enterocolitica biotype 5, was the fourth most common documented cause of death, and the most prevalent infectious cause (Buddle et al. 1988). It was also the most common bacterial cause of caprine enteritis identified in diagnostic laboratories (Vickers 1986). The disease occurs mostly in late autumn or winter and has been linked to the stress of transport, excessive handling of feral goats, nutritional deprivation, winter shearing, cold and wet weather, inadequate shelter, and, in one outbreak, contact with pigs (Orr et al. 1987). Young goats are more often affected, but all ages are at risk. Y. enterocolitica is more frequently identified than Y. pseudotuberculosis (Thompson 1985). Enteric yersiniosis also has been reported in goats in Norway (Krogstad et al. 1972) due to Y. enterocolitica, in Australia due to Y. enterocolitica and Y. pseudotuberculosis (Slee and Button 1990a, b), and in Japan due to Y. pseudotuberculosis (Seimiya et al. 2005).
A cohort study of Yersinia infection in goats in New Zealand indicated a clear seasonal and age-related pattern of infection in goats, with most fecal excretion of pathogenic biotypes of Y. enterocolitica and Y. pseudotuberculosis occurring in the colder winter months and dropping off in summer, while excretion of non-pathogenic or environmental Yersinia spp. (e.g., Yersinia frederiksenii) showed a more persistent temporal pattern. The shedding of pathogenic Yersinia was highest in goats less than 1 year of age, with pathogenic strains rarely isolated from adults (Lanada et al. 2005). It is likely that young goats normally experience subclinical infections due to environmental exposures and develop immunity, but if they are stressed during those infections they may manifest clinical disease (Orr et al. 1990).
Pathogenesis
Infection is usually via the oral route, though in the case of mastitis entry via the teat canal is more likely. The two Yersinia spp. causing yersiniosis are known to contain two plasmid-mediated virulence factors, the V and W antigens, similar to those of Y. pestis. In addition, some Y. pseudotuberculosis serotype III organisms produce an exotoxin that may enhance virulence (Obwolo 1976). Yersinia organisms multiply in the intestine, producing enteritis, but the organisms also produce bacteremia via the portal circulation and lymphatics, leading to internal abscesses, abortions, and acute deaths.
Clinical Findings
The enteric form of yersiniosis occurs most often in kids between 1 and 6 months of age, but all ages and breeds can be affected. In an outbreak in Japan, for instance, 29 of 100 adult lactating does developed diarrhea, but no kids, males, or dry does on the premises became ill (Seimiya et al. 2005). The initial signs were anorexia and a significant drop in milk production followed by depression and a watery diarrhea containing mucus one or two days later that lasted for four to six days. The case fatality rate was 13.7% (4 of 29). There was no fever recorded in any affected animals, but fever has been associated with enteric yers- iniosis in other reports. In young animals, cases of sudden death without diarrhea may occur along with cases of diarrhea. The diarrhea is watery and not bloody. The course of diarrhea may be short, up to several days, and is often fatal. A more prolonged illness with dehydration and weight loss also may be seen. In the abortion form, spontaneous abortions may occur in a herd in conjunction with the birth of weak, term kids that die shortly after birth. The cotyledons on placentas from affected does may be entirely white or have focal white spots on their surfaces (Witte et al. 1985). An abortion rate of 24% was recorded in one affected herd over a period of 10 days and affected does died (Albert 1988).
Both acute and chronic forms of mastitis occur. There is udder swelling and clots in the milk in the acute form and there may be blood in the milk in the chronic form. Induration of the udder may persist for weeks after infection is cleared.
Clinical Pathology and Necropsy
Goats with enteritis may show a pronounced leukocytosis with neutrophilia and a marked left shift (Seimiya et al. 2005). There may also be laboratory evidence of hemoconcentration due to dehydration from diarrhea. The causative organism can be isolated from diarrheic feces, mastitic milk, and uterine discharges. Because the organism may be present in feces as a normal gut inhabitant, serotyping is necessary to confirm the isolate as a likely pathogen. Serologic testing can be used to support a diagnosis of yersiniosis. Goats with diarrhea caused by Y. enter- ocolitica may show marked increases in agglutination titers between sera taken at the time of diarrhea and two to three weeks later (Krogstad 1974).
At necropsy, goats with the enteric form of yersiniosis are thin and often emaciated. The large, multiple, internal abscesses (pseudotuberculosis) characteristic of the disease in laboratory animals are not reported in goats, though occasional, single abscesses have yielded Y. pseudotuberculosis. The most consistent gross finding is enlargement and edema of the mesenteric lymph nodes. Gross lesions in the intestines may be limited to hyperemia or edema of the mucosa or a catarrhal enteritis, but careful inspection of the mucosal surface frequently reveals small focal areas of necrosis visible as small, tan or whitish, circular lesions 1-2 mm in diameter, which may be surrounded by reddish zones of hemorrhage or congestion. Histologically, there are microabscesses in the superficial mucosa or lamina propria of the intestine that contain colonies of Gramnegative bacteria surrounded by neutrophils (Vickers 1986). Gross thickening of the cecum and proximal colon and mucosal ulceration and fibrin plaque formation have also been observed in at least one goat with intestinal yersinio- sis (Slee and Button 1990b). Microabscesses in the small and large intestines are strongly suggestive of Y. enterocol- itica infection (Slee and Button 1990a).
In the abortion form of the disease, the uterus may be hemorrhagic or filled with pus. Goats that die with the abortion form of the disease may also have swollen, edematous mesenteric lymph nodes. Splenomegaly, renal infarcts, and swelling of the intestinal mucosa with edema also have been noted. Yersinia organisms can be cultured from mesenteric nodes and uterine content.
Diagnosis
Because of the zoonotic potential of yersiniosis, definitive diagnosis by bacteriologic or serologic methods should be attempted whenever human exposure is a concern. In the enteric form of the disease, all potential causes of diarrhea must be considered. In the age group most often affected, kids 1-6 months of age, coccidiosis and nematode parasitism are the most likely causes of diarrhea and must be ruled out. Salmonellosis and enterotoxemia are also likely to produce diarrhea in conjunction with sudden deaths. The differential diagnosis for abortion is discussed in Chapter 13, and for mastitis in Chapter 14. Internal abscesses in goats are most likely caused by C. pseudotuberculosis, but yersiniosis, tuberculosis, and melioidosis must also be considered as potential causes.
Treatment
Yersinia are generally susceptible to a wide range of broadspectrum antibiotics. Most reports of successful treatment, however, have involved use of tetracyclines. Tetracycline was effective in halting outbreaks of both the enteric and abortion forms of the disease (Orr et al. 1987; Albert 1988). Antibiotic therapy is most successful when initiated early in the course of disease. In severe diarrhea cases, supportive fluid therapy may improve outcome. Antibiotic sensitivities were conducted on the Y. pseudotuberculosis serotype III isolates associated with diarrhea in lactating goats in Japan. All were sensitive to enrofloxacin, ceftiofur, gentamicin, tetracycline, oxytetracycline, colisitin, and fos- fomycin (Seimiya et al. 2005).
Control
Because the epidemiology of caprine yersiniosis is incompletely understood, control recommendations are largely empirical. In New Zealand, it is suggested that to minimize stress, hair goats normally sheared in winter be well fed before shearing and have good shelter available immediately afterward. It is also recommended that newly captive feral goats not be sheared at all their first winter to avoid compounding their stress (Thompson 1985). Other general recommendations include avoidance of overcrowding and other stressors, provision of adequate nutrition and feeder space, separation of goats from swine, good nematode parasite control, and good rodent and bird control in intensive housing and feeding arrangements. Water supplies should be tested for the presence of pathogenic Yersinia. There are no commercial vaccines available for Y. enterocolitica or Y. pseudotuberculosis in livestock.
Paratuberculosis
Paratuberculosis, also known as Johne's disease, is an economically important infectious disease of domestic and wild ruminant animals, primarily affecting the digestive tract. Infection leads to gradual debilitation and death through a mechanism of digestive dysfunction that is not completely understood.
Because the disease has been studied most intensively in cattle, many aspects of the bovine disease traditionally have been assumed to hold true for goats. However, this is not the case. Diarrhea, which is the cardinal sign in cattle, is an uncommon clinical sign in goats. Caprine paratuberculosis is characterized mainly by chronic, progressive weight loss in adults. Suggestions that the organism causing paratuberculosis in ruminants may be associated with the development of Crohn's disease in humans have spurred interest in recent years in more effective recognition and control of paratuberculosis in cattle, goats, and sheep.
Etiology
Paratuberculosis is caused by the bacterium formerly known as Mycobacterium paratuberculosis (also previously called Mycobacterium johnei). Based on advances in the molecular characterization of the organism that occurred in the 1990s, the organism has since been reclassified as a subspecies of Mycobacterium avium and is referred to as M. avium subsp. paratuberculosis (Map). The characterization and molecular epidemiology of Map have been reviewed (Harris and Barletta 2001). Analysis of Map strains from different geographic locations and different host species using various molecular techniques indicates that there are two main groups of strains. The C, or cattle, strains occur mainly in cattle, but are also the predominant strains in goats. The C strains also occur in deer and, rarely, sheep. The S, or sheep, strains occur mainly in sheep, but also in farmed deer and sometimes goats. The presence of S strains in goats, though less frequent than the C strains, occurs mainly when goats are commingled with infected sheep. Earlier studies suggested that there was a unique goat strain of Map in Norway that had little or no pathogenicity for cattle. However, more recent studies found that there was no genotypic variation between the isolates from cattle and the majority of the Norwegian goat isolates from infected herds and flocks, indicating that the most common strain of Map in Norway is able to infect both cattle and goats (Djonne et al. 2005).
Map is a small mycobacterium (0.5 ? 1 μm) compared with other pathogenic mycobacteria. It demonstrates typical acid fastness with Ziehl-Neelsen stain. In tissues, the organism tends to be found in clumps inside macrophages rather than as individual bacteria. The bacterium is very resistant to environmental degradation and can persist in barnyards and in manure spread on pastures for longer than one year. Shade on pastures prolongs the survival time and there are indications that Map may be genetically capable of dormancy, i.e., being able to enter a viable but non-cultivable state and later reverting to a vegetative form (Whittington et al. 2004). Disinfectants capable of eliminating the organism from the environment include cresylic compounds diluted 1 : 64 and sodium ortho-phenylphenate diluted 1 : 200.
The C strain of Map is a fastidious, slow-growing organism in vitro. Positive cultures are rarely identifiable before 6 weeks and should be held at least 12 weeks for confirmation of negative cultures. The cultivation of field isolates of Map requires supplementation of the culture media with mycobactin, an iron-chelating substance found in other Mycobacterium spp. Mycobactin dependence has long been considered a distinguishing culture characteristic of the paratuberculosis organism. However, it is now known that certain strains of M. avium subsp. avium and M. avium subsp. sylvaticum (formerly known as the wood pigeon mycobacterium) also demonstrate mycobactin dependence and may sometimes infect ruminants. These other mycobactin-dependent species can be distinguished from Map by molecular techniques, based on the fact that Map contains multiple copies of the genetic insertional sequence IS900 while the other species contain the genetic inser- tional sequences IS901 (M. avium subsp. avium) or IS902 (M. avium subsp. sylvaticum).
Until recently, the S strains of Map could not be grown using the standard culture techniques successfully employed for cultivation of the C strains. Reliable growth was found to be possible by use of liquid modified BACTEC 12B medium, but this use of radiometric liquid media has now been replaced with non-radiometric liquid medium M7H9C, which also detects both C and S strains (Whittington et al. 2013). Solid media (modified Middlebrook 7H10 and 7H11 agars) can also support growth, but may be less sensitive (Whittington et al. 1999).
The interspecies infectivity of Map was established in 1913 when clinical paratuberculosis was produced in a goat with a bovine-derived inoculum (Twort and Ingram 1913). In general, when different ruminant species are managed together on a single farm premises and one species is infected, the risk of cross-infectivity is considerable. The development of paratuberculosis in feral goats in New Zealand after introduction into paddocks with known paratuberculous cattle has been documented (Ris et al. 1988).
Epidemiology
Paratuberculosis was first definitively described in cattle in 1895 (Johne and Frothingham 1895) and in goats in 1916 (McFadyean and Sheather 1916). It has traditionally been considered a disease of temperate regions, with sporadic occurrence in tropical environments, primarily as a result of importation of infected livestock from endemic areas. Today, paratuberculosis is generally considered a disease of worldwide distribution. However, accurate details on the geographic distribution of caprine paratuberculosis are difficult to ascertain, because statistics on goat disease are often reported together with those on sheep by governments and international agencies.
Caprine paratuberculosis has been specifically described in numerous countries, representing all continents but Antarctica. These countries include the Sudan (Chaudhari et al. 1964) in Africa; India (Rajan et al. 1976), Nepal (Singh et al. 2007), and Korea (Lee et al. 2006) in Asia; Turkey (Alibasoglu et al. 1973), and Israel (Shimshony and Bar Moshe 1972) in the Middle East; Cyprus (Polydorou 1984), France (Yalcin and Des Francs 1970), Greece (Xenos et al. 1984), Norway (Holmboe and Slagsvold 1934), Spain (Vizcaino et al. 1984), and Switzerland (Tontis and Konig 1978) in Europe; Canada (Morin 1982; Moser 1982), the United States (West et al. 1979; Sherman and Gezon 1980; Ullrich et al. 1982), and Mexico (Ramirez et al. 1983; Estevez-Denaives et al. 2007) in North America; Chile (Kruze et al. 2007) in South America; and Australia (Lenghaus et al. 1977; Straube and McGregor 1982) and New Zealand (Thompson 1986) in Australasia.
The prevalence of caprine paratuberculosis is not well documented and presumably it may vary widely between countries because of local management conditions, infectivity of Map, and other factors. The prevalence is likely to be higher in intensively managed goats than in extensively managed goats. For example, in Norway, where goats are kept intensively for dairying and a goat-specific strain of the organism is recognized, nationwide prevalence was as high as 53% before the introduction of a vaccination program. In India, with generally smaller herds and more extensive management, a 13-year necropsy study indicated a prevalence of only 5.2% (Kumar et al. 1988).
A significant driver for a better understanding of the epidemiology of paratuberculosis has been the suggestion of a putative association between M. avium subsp. paratuberculosis and Crohn's disease, a granulomatous enteritis of humans. Paratuberculosis was proposed as a possible model for the study of Crohn's disease in 1972 because of the clinical and histopathologic similarities of the two conditions, but at that time there was no suggestion that the Map organism itself might be involved in the pathogenesis of the human disease (Patterson and Allen 1972). However, in 1984 the Map organism was identified in tissues of four Crohn's disease patients (Chiodini et al. 1984), suggesting an etiologic role for the organism. This also suggested that paratuberculosis might be a zoonotic disease, potentially transferable by contact with infected livestock or by consumption of food products from those animals. Though no causal relationship was established, a recent study conducted in the Fars province of Iran, where small ruminant production is prominent, identified Map in the biopsies of 47% of human Crohn's disease patients sampled and in 70% of biopsies taken from healthy goats sampled at slaughterhouses in the province (Zarei-Kordshouli et al. 2019).
Considerable research has been carried out on the possible role of Map in the pathogenesis of Crohn's disease and that literature has been reviewed (Hermon-Taylor et al. 2000; National Research Council of the National Academies 2003). A body of research has also evolved relative to the potential for zoonotic transmission from infected animals, meat, milk, or milk products, and this also has been reviewed (Grant et al. 2001; Grant 2006). Concerns about the risk of dairy products were heightened when it was demonstrated in the United Kingdom that Map in milk can survive standard high-temperature-short-time pasteurization of 72 °C (161.6 °F) for 15 seconds (Grant et al. 1999). This prompted regulators and processors to extend the pasteurization time to 25 seconds. Still, Map could be identified in containers of pasteurized milk offered for sale at the retail level (Grant et al. 2001; O'Reilly et al. 2004). A recent study suggests that the persistence of Map in milk following pasteurization is a result of the bacteria residing in somatic cells passed from the udder being protected from heat inactivation and is not due to fecal contamination of the milk (Gerrard et al. 2018). A comprehensive study by the National Research Council published in 2003 concluded that “there remains insufficient evidence to prove or disprove that Mycobacterium avium subsp. paratuberculosis is a cause of some or all cases of Crohn's disease in humans” (National Research Council of the National Academies 2003). However, it was also concluded that a causal link between paratuberculosis and Crohn's is plausible, and a new research approach is needed to definitively establish or disprove that relationship. Nevertheless, there are members of the human medical community who believe quite strongly that sufficient evidence exists to make that link and they call for a strong and immediate public health response (Hermon-Taylor and Bull 2002). Others believe that the evidence for involvement of Map in Crohn's disease remains unclear (Eckburg and Relman 2007; Behr and Kapur 2008).
In the meantime, public perceptions have spurred regulators and producers to take the control of paratuberculosis in cattle, sheep, and goats far more seriously than in decades past. In a recent survey, a majority of topic specialists agreed that Map is likely a risk to human health and that it is a high- or medium-priority agri-food public health issue. The most highly ranked sources of Map for humans in that survey were considered to be dairy products and ruminants, but meat, produce, and water may also be contributing sources (Waddell et al. 2016). In a study of ewes from a sheep farm in New Zealand with a clinical history of paratuberculosis, Map was identified by radiometric culture in skeletal muscle of 15 and the blood of 13 of 18 ewes confirmed as infected with Map on the basis of histopathologic evidence typical of paratuberculosis within the ileum and adjacent lymph nodes (Smith et al. 2011).
Pathogenesis
The primary mode of transmission of paratuberculosis is fecal-oral, with the organism shed in the manure of infected adults and ingested by susceptible young stock, particularly when animals are overcrowded and sanitation is poor. An age-related resistance to infection is well known to exist in cattle, and this appears to hold true for goats as well (Levi 1948). Neonates are considered most susceptible to new infections, particularly when their own dam is an active fecal shedder and the kid is allowed to remain with the doe. However, age-related resistance is not absolute, and it is probable that adult animals may remain at risk for new infection if kept in overcrowded, heavily contaminated environments.
In utero infection of the fetus is considered a less common form of transmission, but is documented in cattle (McQueen and Russell 1979) and sheep (Tamarin and Landau 1961). The importance of in utero transmission in the goat remains unclear. The organism has been recovered from the uterus and fetal organs of some experimentally infected goats (Goudswaard 1971) and from the fetus of a naturally infected Pygmy goat with advanced clinical paratuberculosis (Alinovi et al. 2009). As a practical matter, currently accepted disease control practices assume the possibility of in utero transmission and recommend the culling of offspring of confirmed infected does.
As with cattle and sheep, goats are most likely infected at an early age by ingestion of the organism. Following ingestion, the organism localizes in the mucosa of the small intestine and associated lymph nodes. The organism is transported across the intestinal mucosa by M cells associated with the Peyer's patches or by enterocytes (SigurdSardottir et al. 2005). While some exposed individuals may develop resistance to chronic infection, many infected goats subsequently carry the infection in a dormant state in the Peyer's patches of the intestine and the mesenteric lymph nodes for a variable period into adulthood. At some point, triggered by stress or other ill-defined factors, some infected animals begin to shed the organism in the feces. They may begin to show clinical signs concurrently or at a later time. These signs include diarrhea and weight loss in cattle, and progressive weight loss without diarrhea in sheep and goats.
It is not known why diarrhea is the distinguishing characteristic in bovine paratuberculosis, but an infrequent finding in the caprine and ovine forms of the disease. Because small ruminants normally produce a drier feces than the cow, it may require a more severe derangement of colonic water resorption capacity in the goat or sheep to produce evidence of clinical diarrhea than in the cow.
In general, small ruminants with paratuberculosis tend to have less severe lesions of granulomatous enteritis than affected cattle. There have been a number of studies of experimental paratuberculosis infection in goats aimed at gaining a better understanding of the pathogenesis of the disease at a cellular level and the associated cell-mediated and humoral immune responses (Storset et al. 2001; Valheim et al. 2002, 2004; SigurdSardottir et al. 2005; Munjal et al. 2005; Stewart et al. 2006). A standard experimental challenge model for caprine paratuberculosis has been proposed (Hines et al. 2007).
There is experimental evidence that bacteremia occurs in goats with clinical paratuberculosis based on isolation of the organism in blood cultures and numerous tissues at necropsy, including the udder and uterus (Goudswaard 1971). This suggests that offspring born to clinically infected does have a very high likelihood of infection through the birth process or when suckling, if not already infected as fetuses.
Because of the prolonged dormancy of infection, the persistence of the organism in the environment, and the endemic nature of the disease, all goats in a known infected herd must be considered at risk of infection. Goats in an infected herd may fall into one of four categories: resistant or non-infected individuals, infected non-shedders, sub- clinically infected inapparent shedders, and clinically affected apparent shedders. Only individuals in the last group can be identified as abnormal on physical examination, although a definitive diagnosis can never be made on physical examination alone. The subclinical infection rate in an infected herd is likely to be much higher than the rate of clinically apparent cases.
Clinical Findings
Overt clinical disease rarely occurs before 1 year of age and is most common in goats 2 and 3 yeans of age. Clinical disease is often triggered by some episode of stress, such as parturition or recent introduction into a new herd. Affected individuals begin a course of pro - gressive weight loss, which may extend from weeks to months and can lead to dramatic emaciation (Figure 10.6). Appetite may remain intact initially, but decrease later on, and the animal becomes increasingly lethargic and depressed. A rough haircoat and flaky skin are common. Animals in the advanced state of disease may become immunologically anergic. Advanced cases eventually succumb to debilitation, inanition, exposure, or secondary infections.
Unlike cattle, goats rarely show persistent watery diarrhea, except possibly in the terminal stages of disease. When it occurs, diarrhea may actually be due to concurrent parasitism, although sometimes it may be directly because of paratuberculosis. Soft stool, like dog feces in appearance, may occur intermittently through the course of disease, but pelleted feces are the rule.

Figure 10.6 Adult Nubian buck with advanced clinical paratuberculosis. Note severe emaciation. Source: Courtesy of Dr. David M. Sherman.
As the disease progresses a moderate anemia of chronic infection may develop and clinical evidence of hypoalbu- minemia such as intermandibular edema may be seen. The non-specific nature of these clinical signs makes a definitive diagnosis of paratuberculosis by clinical examination impossible.
Clinical Pathology and Necropsy
Anemia and hypoalbuminemia are likely to be present in advanced clinical cases, but are not specific for paratuberculosis. Hypergammaglobulinemia and hypocalcemia have also been reported in affected goats (Schroeder et al. 2001). Definitive diagnosis of paratuberculosis requires bacteriologic or serologic testing and/or histopathologic examination of tissues.
Infection with paratuberculosis slowly evolves in such a way that no single diagnostic test can possibly detect all infected animals at any given time in the course of the disease. The earliest detectable reactions to develop involve the cell-mediated immune (CMI) response. The humoral immune response, with the production of circulating antibodies, occurs after the CMI response, later in the course of subclinical infection. The detection of infection by culture of organisms from the feces can only occur in animals that are actively shedding the bacteria, and the onset of shedding is variable relative to the humoral immune response. Some subclinically infected animals with a detectable antibody response may remain non-shedders for a significant period of their infection. One experimental study tracked the progression of the goat host response to Map infection. The CMI response, as measured by the lymphocyte proliferation test, was detectable at 60 dpi; detectable fecal shedding of Map began between 150 and 180 dpi; and antibody response, measured by ELISA and AGID, became detectable at 180 dpi (Munjal et al. 2005).
During clinical disease, fecal culture and tests of cell- mediated and humoral immunity can be employed for diagnosis, but in the advanced stages of clinical disease anergy may occur in some individuals and cell-mediated and humoral immune responses may be lost, yielding falsenegative results. Furthermore, even when tests are applied at the appropriate times, there is no single bacteriologic or serologic test with sufficient sensitivity to identify all clinical and subclinical cases of caprine paratuberculosis or with sufficient specificity to avoid some false-positive results. Variations in fecal shedding can also lead to falsenegative culture results. For these reasons, more than one type of test needs to be applied for implementation of herd control programs that include testing and culling of infected animals.
Traditionally, the CMI response has been assessed by in vivo intradermal skin test using johnin purified protein derivative (PPD-J) or the intravenous johnin test. These in vivo assays have largely been replaced by newer techniques involving the identification of cytokines released by immune cells, such as the production of gamma interferon (IFNγ) produced in vitro by stimulation of peripheral blood mononuclear cells with mycobacterial antigens, and then measured using a sandwich enzyme immunoassay (Rothel et al. 1990). The IFNγ assay was used to monitor the progression of paratuberculosis in a naturally infected herd of Pygmy goats along with fecal culture, ELISA, and AGID. The IFNγ assay produced both false-positive (1 of 3) and false-negative (3 of 10) results in live animals, based on the true infection status determined subsequently by necropsy and culture of Map from tissues (Manning et al. 2003). In experimentally challenged goats, CMI was measured by IFNγ assay, but also by interleukin 2 receptor expression and by lymphocyte proliferation test (Storset et al. 2001). CMI responses were detectable by nine weeks post inoculation and persisted with variation through the two years of the study, though responses were stronger in the first year.
The tests currently in use for identification of antibody responses in goats are the complement fixation (CF) test, the AGID test, and the ELISA test. In general, when used in goats, these tests have a sensitivity in the range of 85-100% when applied to clinical cases, but only 20-50% when applied to subclinical cases (Stehman 2000).
The CF test is probably the least commonly employed, but remains in use because a number of countries require a negative CF test for importation of ruminant livestock. One problem with the CF test is that anticomplementary activity with goat serum may confound interpretation of the test, yielding neither clear positive nor negative results (Stehman 1996). Specificity may also be reduced in the CF test due to cross-reactions with other mycobacteria or C. pseudotuberculosis, as discussed further below in relation to the ELISA test.
The AGID test is very effective in confirming suspect clinical cases of caprine Johne's disease, with a sensitivity of 86% or greater, and it provides results within 24-48 hours. It can also identify subclinical shedders with moderate reliability, but sensitivity falls dramatically when the test is applied to subclinically infected non-shedders. AGID tests performed as well as fecal culture when both tests were used for several years in a large, naturally infected goat herd (Sherman and Gezon 1980). Precipitating antibody was detectable in infected goats at approximately the same time that the animals began to shed the organism in their feces. Similar results were reported independently in a large goat herd studied for three years in England, where fecal culture and the AGID test proved more useful than intradermal and CF tests for detecting infected individuals (Thomas 1983). Nevertheless, false-negative AGID test results do occur in subclinically infected goats and a single negative test should not be presumed to indicate noninfected status (Ramirez Casillas et al. 1984).
The application of ELISA tests for the diagnosis of paratuberculosis emerged in the 1990s and since then a number of studies have reported its use specifically in goats (Milner et al. 1989; Molina et al. 1991; Molina Caballero et al. 1993; Burnside and Rowley 1994; Rajukumar et al. 2001; Whittington et al. 2003; Dimareli-Malli et al. 2004; Munjal et al. 2004; Gumber et al. 2006). Differences in performance are noted, often as a result of the type of antigens that are employed in the assay. In general, these ELISA tests have sensitivity in the range of 54-88%; they are more reliable for confirmation of clinical cases than for detection of sub- clinical cases. In experimentally infected goats, ELISA and AGID tests were found to be 100% sensitive from 180 and 210 dpi onward, respectively (Munjal et al. 2005).
The specificity of ELISA tests may suffer because of the occurrence of cross-reactions. Map is known to share common antigens with other Mycobacterium spp., Nocardia spp., and Corynebacterium spp. Corynebacterial infections may produce cross-reactivity in serologic tests used to diagnose Map infection (Ridell 1977). This is of particular concern in goats, because caseous lymphadenitis caused by C. pseudotuberculosis is a common disease in the species. The problem of cross-reaction occurs mainly with ELISA and CF tests, while the AGID test is not influenced by antibodies to C. pseudotuberculosis (Van Metre et al. 2000). In one study comparing specificity of two paratuberculosis ELISA kits and the AGID test, 344 goats with known exposure to C. pseudotuberculosis but with negative fecal cultures for Map were tested. The AGID gave no false-positive results. One ELISA kit gave five false-positive results (1.5%), while the second kit gave 89 false positive results (25.9%) (Manning et al. 2007). Clearly, if the infection status of a herd is not known relative to paratuberculosis and caseous lymphadenitis, the ELISA test would not be a wise choice for screening the herd to determine the prevalence of paratuberculosis. The use of absorbed ELISA assays wherein serum is pretreated with Mycobacterium phlei to reduce cross-reactivity has improved the specificity of the tests.
The comparative performance of a commercial ELISA has been evaluated for goat milk and serum from the same animals. Sensitivity of the milk ELISA was lower than for the serum ELISA, but the specificity was higher than that of the serum ELISA (Salgado et al. 2005, 2007). More recently, ELISA testing on goat serum and goat milk are reported to have comparable sensitivities and specificities, suggesting that ELISA testing using milk samples in dairy goat paratuberculosis control programs may be advantageous, because milk sampling is non-invasive, rapid, and easy compared to serum sampling (Angelidou et al. 2014).
There have been some notable advancements in techniques for identification of the organism in the feces or tissues of infected animals. Conventional fecal cultures on solid media grow slowly. Positive cultures can take 6 weeks or more to develop, and culture tubes have to be held for a minimum of 12 weeks to ensure that they are negative. The use of liquid media has allowed for faster cultivation. Radiometric techniques for culture of mycobacteria on liquid media have been adapted for the growth of Map and provide more rapid results with equal or greater sensitivity than conventional culture on solid media (Collins et al. 1990; Sockett et al. 1992; Eamens et al. 2000). The use of radiometric liquid medium also enabled laboratories to culture the S strain of Map from sheep (Whittington et al. 1999). Since 2012, however, the radiometric technique has been discontinued and replaced with a nonradioactive liquid medium known as MH9C, with similar sensitivity and the potential for cultivation of C and S strains of Map, both of which may infect goats (Whittington et al. 2013).
Molecular probes and PCR techniques have been developed to confirm the presence of Map-specific genetic material in feces or tissue samples, in an effort to bypass the need for culture. However, inhibitors in feces and inefficient DNA release from clinical samples initially hampered the general application of these techniques. More recently, sensitivities of IS900-based PCR assays for the detection of Map from both fecal and tissue samples have improved, and enhanced DNA extraction techniques allow the use of qPCR assays using primers that avoid detection of environmental bacteria (Kawaji et al. 2011). Issues regarding the use of PCR in the diagnosis of paratuberculosis have been reviewed (Bolske and Herthnek 2010).
Expense is an issue in herdwide testing for paratuberculosis and may be an obstacle in getting herd owners to undertake testing either to establish herd prevalence or to follow through with test and cull programs for control. One approach to reducing costs has been the use of pooled fecal cultures to determine a herd's infection status. This is done by combining samples from groups of animals of the same age for testing. The sensitivity of this method depends on the prevalence of infection in the herd and the intensity of shedding among herd members. In a report from Australia on the use of pooled samples for culture of Map in goats, pools of 25 samples were considered to be suitable for producing meaningful test results based on the conditions prevailing in that study (Eamens et al. 2007).
Because clinical caprine paratuberculosis is invariably a terminal disease, necropsy provides an important opportunity for definitive diagnosis given the various challenges associated with diagnosis in live animals. Gross lesions are more variable in occurrence in the goat compared with cattle. Therefore, histologic examination of tissues and bacterial culture and PCR of selected organs also should be performed.
The gross and microscopic lesions of caprine paratuberculosis are well described (Levi 1948; Harding 1957; Nakamatsu et al. 1968; Fodstad and Gunnarsson 1979). The prominent, accordion-like, corrugated thickening of the intestinal wall and mucosa commonly seen in cattle is not often observed in goats. When gross lesions are present in small ruminants, they usually present as focal or diffuse thickening or edema of the ileum, cecum, or spiral colon. Adjacent lymph nodes may be enlarged and edematous, and caseation of nodes with focal calcification occurs more often in goats than in cattle. Emaciation is a constant finding, although persistence of abundant mesenteric fat deposits is not unusual in goats, even when the general body condition is one of emaciation. Calcification of the aorta often occurs in goats with prolonged paratuberculosis, though the pathogenesis is unknown. Evaluation of live goats using transabdominal ultrasonography showed a strong correlation between the detection of enlarged mesenteric lymph nodes and confirmation of paratuberculosis at necropsy. Thickening of the small intestinal mucosa was also detected by ultrasound, but did not correlate as well as mesenteric lymph node enlargement at necropsy in confirmed paratuberculous goats (Tharwat et al. 2012)
Histologically, paratuberculosis produces characteristic granulomatous lesions in the intestinal tract, lymph nodes, and possibly liver. At a minimum, tissues submitted for microscopic examination should include a section of the ileocecal junction and adjacent ileocecal lymph node, ileum, spiral colon, and another mesenteric node. Where available, PCR can be applied to paraffin-embedded tissues to detect the IS900 insertional sequence characteristic of Map. Otherwise, acid-fast Ziehl-Neelsen or immunohistochemical staining should be requested on these samples when paratuberculosis is suspected. Bacterial culture of fresh or frozen tissues is also possible. In areas where both tuberculosis and paratuberculosis occur in goats, microbiologic diagnosis is imperative because necropsy lesions may be similar.
Diagnosis
The differential diagnosis of paratuberculosis in goats is essentially the differential diagnosis of chronic weight loss. The causes of this common presenting sign in goats are covered in detail in Chapter 15.
Treatment
There are no known effective treatments for the elimination of Map infection and none is specifically approved for the treatment of paratuberculosis. Various antimycobacterial drugs have been used with limited effect in goats. Attempted unsuccessful therapies have included isoniazid, isoniazid plus rifampin, isoniazid plus ethambutol, or all three drugs together (Gezon et al. 1988). However, some treatment regimens have alleviated clinical signs in individual cases as long as therapy is continued. Nevertheless, the organism can still be isolated from cultures prepared from tissues at necropsy. One such regimen is daily streptomycin sulfate (0.5 g intramuscularly) plus isoniazid (25 mg by mouth) plus sodium aminosalicylate (850 mg by mouth) for six months (Zahinuddin and Sinha 1984). A second regimen is daily dihydrostreptomycin (0.5 g intramuscularly) plus rifampin (300 mg by mouth) and isoniazid (300 mg by mouth) twice daily (Slocombe 1982). Long-term administration of monensin has been reported to have some therapeutic effect in cattle, reducing the severity of paratuberculous lesions in the digestive tract (Brumbaugh et al. 2000) and reducing fecal shedding of Map in calves (Whitlock et al. 2005), but no trials in goats have been reported. Treatment has little economic application in goats, except for the possibility of keeping known infected, high-quality breeding does in good health long enough to achieve embryo transfers or to prolong the life of pet goats. However, in light of the putative connection between paratuberculosis and Crohn's disease, owners of pet goats should be made aware of that link before deciding to treat and maintain a known infected goat as a pet.
Control
Interest in the control of paratuberculosis has increased in recent years, mainly for two reasons. The first is increased understanding of the economic losses associated with the disease, particularly in dairy herds. In dairy cattle, losses are associated with decreased milk production, decreased milk fat and protein, premature culling, reduced slaughter weight at culling, decreased fertility, reduced feed efficiency, and increased incidence of mastitis (Constable et al. 2017). Losses have been estimated to be as high as $200/cow/year in high prevalence herds. While similar types of loss could be expected in goat dairies, reports on the actual economic impact of paratuberculosis in infected dairy goat herds are scarce. Control of paratuberculosis in goats and sheep has been reviewed (Juste and Perez 2011; Windsor 2015).
The second major stimulus for control of paratuberculosis is the possible zoonotic potential of the disease relative to Crohn's disease in humans, which has caused concern among public health officials, regulatory veterinary agencies, and livestock producers about public perception of milk and meat as wholesome foods and with regard to international trade in goats and goat products. As a result, numerous countries have initiated or expanded their paratuberculosis control programs in recent years. Most programs focus on cattle, but some may include sheep and goats. Readers should contact their own regulatory veterinary authorities to determine the status of goats in existing paratuberculosis control programs. In the United States, for example, there is a Voluntary Bovine Johne's Disease Control Program that focuses on control of the disease in cattle. It is managed by the US Department of Agriculture, Animal and Plant Health Inspection Service (USDA APHIS). Johne's disease programs for sheep and goats, where they exist, are administered through the individual states rather than at the national level. Veterinarians and goat owners should contact the state veterinarian's office in their own state to determine if paratuberculosis in goats is considered a reportable disease, if there are any paratuberculosis testing requirements for movement of goats in or out of the state, and if there is any sort of caprine paratuberculosis control program in place.
Paratuberculosis is almost always a herd problem rather than an individual animal problem, unless the individual goat diagnosed with paratuberculosis was recently introduced into the herd. When a clinical case is diagnosed in an animal long present in an established herd, the herd almost inevitably includes additional subclinical shedders and infected, non-shedding goats. These subclinical infections are likely to be present in far greater numbers than the clinical cases, often in the range of one to five subclinical cases for each clinical case, depending on herd size, how long the herd has been in existence, and various management factors that affect the level of bacterial contamination on the farm. Environmental factors, including soil characteristics such as pH and iron content, may also contribute to the persistence of the organism (Elliott et al. 2015).
There are three basic elements to consider in developing a strategy for eliminating paratuberculosis from infected herds: testing and removing infected individuals from the herd; reducing the rate of new infections in susceptible kids through improved biosecurity and modified kid-rearing techniques; and vaccinating to increase host resistance to new infections. It is unlikely that any one of these components of the control program taken alone will effectively eliminate paratuberculosis, but the emphasis on each component in the overall control program will vary with local conditions. On the national level, caprine paratuberculosis has been controlled in Norway largely through vaccination (Saxegaard and Fodstad 1985) and the economic benefits for goat owners to participate in national disease control programs against paratuberculosis, caseous lymphadenitis, and CAE have been assessed (Nagel-Alne et al. 2014).
At the herd level, elimination of paratuberculosis in a large commercial goat herd in the United States used for antibody production was achieved without vaccination by testing and culling in conjunction with altering management procedures (Gezon et al. 1988). Total eradication through testing and culling and improved biosecurity without vaccination has also been reported in a herd of dairy goats used for production of biopharmaceuticals (Gavin et al. 2018). In another report involving a regular dairy goat herd, infection in adult goats was significantly reduced over a two-year period by serologic testing with ELISA and AGID, culling of positive animals, and improving herd management practices (Hutchinson et al. 2004). Note that the two reported instances of totally elimination of paratuberculosis in goat herds occurred in specialized herds maintained for the biomedical industry. In commercial dairy herds, where fewer resources may be available, total elimination of infection may be more aspirational than achievable, but efforts to control the disease and reduce the burden of infection should nevertheless be considered, as improvements can occur.
The best techniques currently available for identifying infected goats are the bacterial culture of feces and the ELISA or AGID tests. Because these tests do not reliably identify all infected non-shedders, testing must be carried out at frequent intervals to assure that infected non- shedders are rapidly identified as they convert to subclini- cal, inapparent shedder status. Testing at intervals of at most six months is recommended. The ability to conduct ELISA tests on milk samples can reduce the time and cost associated with sample taking. In aggressive eradication programs, all offspring of positive test animals are culled in addition to the positive animal itself. This reflects the belief that kids of infected dams are also very likely to be infected. As bacterial culture of feces can be quite costly on a herdwide basis, pooling of fecal samples can be used on initial herd assessments to determine the overall prevalence of the disease (Windsor 2015).
Improved management and biosecurity help reduce the incidence of new infections in young stock. In intensive dairy operations, kidding pens should be cleaned or rebedded between kiddings. Kids should be taken immediately from the does at parturition, before they are licked, and raised in separate facilities on heat-treated colostrum and pasteurized milk or milk replacer. Kids should not commingle again with adult animals until they themselves have kidded. Adult animals should not be overcrowded, manure should be removed frequently from the pens, and feed and water should be provided in a manner that does not allow for fecal contamination. If manure is spread on pastures, goats should not be allowed to graze those pastures for at least one year. The use of pastures in general may be problematic, as it has become clear that wildlife reservoirs exist for Map that may contaminate pastures, even when paratuberculous livestock have been kept off those pastures. An association between Map-infected wild rabbits on farms in Scotland and paratuberculosis in cattle on those farms has been reported (Greig et al. 1999). Wild ruminants are often seen in pastures in North America, and Map has been isolated from a number of species, including whitetail deer, mule deer, bison, and elk (Harris and Barletta 2001).
Management recommendations for extensively managed goat herds are more difficult to make. At the least, does should be turned out on clean, uncontaminated pastures before kidding so they can spread out to kid and minimize cross-contamination. Herdwide testing by culture or PCR of feces or AGID tests to identify infected dams should be performed before the kidding season begins. Infected dams should be either culled or segregated from the non-infected animals before kidding.
Vaccination of goats for paratuberculosis in Norway has been extremely effective in reducing the national prevalence of caprine paratuberculosis (Saxegaard and Fodstad 1985). The vaccine employed initially was a live, attenuated, adjuvanted vaccine containing two strains of Map, and is given to goat kids between 2 and 4 weeks of age. Postmortem evaluation of vaccinated and unvaccinated goats over a 15-year period indicated that the infection prevalence was reduced from 53% in 1966 to 1% in 1982, primarily through the use of vaccination. The majority of infected goats identified in 1982 were not vaccinated.
More recently, it was reported that a killed vaccine (Gudair®, CZ Veterinaria, Pontevedra, Spain) administered to adult goats in a known infected herd was very useful in the control of paratuberculosis. Half the goats were vaccinated, the other half left unvaccinated, and all goats culled for any cause were necropsied to determine the presence of paratuberculosis. By the end of two years, three times as many goats had been culled from the unvaccinated group as the vaccinated group, and three times as many of the cases culled in the unvaccinated group were cases of paratuberculosis (Corpa et al. 2000).
This killed vaccine is now registered in or available for use by special permit in Australia, Cyprus, Germany, Greece, India, the Netherlands, New Zealand, Norway, Spain, South Africa, the United Arab Emirates, and the United Kingdom (A. Colmeiro, Manager of Business Development, CZ Veterinaria, personal communication, February 2008). Despite this widening availability of the killed vaccine for goats, there is little published information regarding the impact of vaccination in goats as compared to sheep and cattle. In 24 trials reported in 9 articles, positive effects of vaccination in goats have been reported in 92% of cases (Juste and Perez 2011).
In those places where vaccination can be included in control programs, it is recommended that all kids be vaccinated between 4 and 16 weeks of age and that subsequent kid crops be vaccinated at the same age, until all animals in a farm have been vaccinated and all non-vaccinated animals have been removed (Lacasta et al. 2015; Windsor 2015). It should be made clear to herd owners that vaccination does not eliminate infection and that vaccination alone without improved biosecurity, kid management, and testing and culling will not eradicate the disease. The benefits of vaccination include a notable decrease in the number of clinically affected animals over time, a decrease in mortality, a gradual decrease in the prevalence of infection, and a notable decrease in the fecal shedding of Map.
Figure 10.7 Persistent granulomatous reaction (arrows) at site of vaccination for paratuberculosis with live vaccine. Source: Courtesy of Dr. David M. Sherman.
There are three potential drawbacks to the use of vaccines containing Map. The first is the appearance of a granulomatous nodule at the site of vaccination (Figure 10.7). The severity of the nodule can be minimized by proper subcutaneous injection and avoidance of intramuscular injection. The second is the fact that animals vaccinated against Map may develop cross-reactivity to standard tuberculosis tests, thus complicating the testing procedures for regulatory programs designed to control tuberculosis. Implementation of paratuberculosis vaccination, then, may be restricted by regulatory agencies in countries where tuberculosis is monitored or still endemic. The third drawback is that accidental injection of the vaccine into people, notably the vaccinator and animal handler, can produce a serious, persistent granuloma at the accidental injection site. Both live and killed vaccines carry this risk. Efforts are being made to develop new novel Map vaccines that address some of these issues (Bannantine et al. 2014; Thukral et al. 2020).