Specific Diseases of the Digestive System Viral Diseases
Peste des Petits Ruminants
This important virus disease of goats and sheep is endemic in much of Africa, the Middle East, and Asia, and is not known to occur in the Western Hemisphere.
It is a highly contagious transboundary disease that has continued to spread to new countries in recent years. It can cause high morbidity and mortality in naive flocks and herds and is seen as a major constraint on small ruminant production in many developing nations. The World Organisation for Animal Health (OIE) and the Food and Agriculture Organization of the United Nations (FAO) launched a PPR global eradication campaign in 2015, with the goal of eliminating the disease by 2030.Etiology
PPR is caused by a morbillivirus in the family Paramyxoviridae. The morbillivirus genus also includes the canine distemper, measles, and rinderpest viruses and the related viruses of cetacean and phocine marine mammals. These RNA viruses are enveloped, contain RNA in tightly coiled helical nucleocapsids, and are structurally, physico- chemically, and antigenically similar. Historically, the virus causing PPR has been called the peste des petits ruminants virus (PPRV), but the International Committee on Taxonomy of Viruses approved a name change to small ruminant morbillivirus (SRMV) in 2019 (Amarasinghe et al. 2019).
There is no evidence of antigenic differences among various field isolates of SRMV; all belong to one serotype. However, phylogenetic analysis based on the partial genome sequence of either the viral N or F gene indicates that there are four distinct lineages for the SRMV (Banyard et al. 2010; Muniraju et al. 2014). Historically, lineages I and II were found in West Africa and lineage III mainly in eastern Africa, while lineage IV was found mainly in the Middle East and Asia. However, lineage IV is now found throughout PPR-endemic areas and is frequently associated with outbreaks in Africa (Parida et al.
2015).SRMV can be cultivated in sheep or goat kidney cell, Vero cell, and embryonal or neonatal testicular cell cultures. A cytopathic effect (CPE) with formation of syncytia and clock-faced giant cells is observed. Intracytoplasmic and intranuclear inclusions occur.
Epidemiology
PPR was first described as a disease mimicking rinderpest in sheep and goats in the Ivory Coast in 1942 (Gargadennec and Lalanne 1942). Serologic cross-reactions with rinderpest virus confused early recognition of the disease as a distinct entity of separate etiology. By the late 1970s, experimental infections, virologic studies, and serologic studies established that PPR was a disease distinct from rinderpest and that the small ruminant disease syndromes of West Africa known as Kata and stomatitis pneumoenteritis complex were in fact PPR (Rowland et al. 1971; Hamdy et al. 1976; Gibbs et al. 1979).
From the 1970s PPR has been identified progressively further eastward in sub-Saharan Africa (Roeder et al. 1994), the Middle East, and South Asia. An epizootic of highly virulent PPR developed in South Asia in the early 1990s and spread widely between Bangladesh, Turkey, and the Middle East. The disease is still actively spreading. The People's Republic of China reported PPR to the OIE for the first time in July 2007. The outbreak occurred in Tibet. PPR was reported for the first time in the countries of Georgia in 2016, Mongolia in 2016, and Bulgaria in 2018. The Bulgarian outbreak represented the first incursion of PPR into the European Union. North Africa has also experienced PPR, with outbreaks occurring in Tunisia, Morocco, and Algeria between 2008 and 2016. By 2019, over 70 countries had reported the disease to the OIE. Thailand reported its first outbreak of PPR in February 2021 following an importation of goats from West Africa. This was the first report of the disease in Southeast Asia.
The principal hosts of PPR are sheep and goats. Breed variation in susceptibility among goats has been suggested.
However, management and climatic factors may be con- founders in this analysis (Ezeokoli et al. 1986). For example, different rates of breeding may affect population susceptibility, as the more prolific breeds may have a higher proportion of younger, more susceptible animals in the population. The disease is considered to be a major constraint in the development of small ruminant productivity among smallholders in developing countries.Virus replication occurs in cattle and pigs, and serumneutralizing antibodies may occur. Neither species, however, is considered to play an active role in disease transmission. Rats can be subclinically infected, but do not appear to transmit the virus to other rats or goats (Komolafe et al. 1987). Seroprevalence studies in Africa and the Middle East indicate that camels are susceptible to SRMV and clinical cases in camels with laboratory confirmation of viral antigen have been reported from Iran (Zakian et al. 2016). The role of camels in the transmission of SRMV to sheep and goats, if any, has not been established.
Some wild ruminants can be infected with PPR and experience clinical disease, but the role of wild ruminant populations in the epidemiology of the disease remains unclear. The whitetail deer of North America is readily infected experimentally (Hamdy and Dardiri 1976). An outbreak of PPR in a zoological park in the United Arab Emirates resulted in peracute deaths in gazelles, ibex, and gemsbok. Goats grazing on the zoo perimeter were believed to be the source of infection (Furley et al. 1987). In 2002, a PPR outbreak was confirmed in semi-free-ranging Dorcas gazelles and Thomson's gazelles in Saudi Arabia, with case fatality rates of 100% (Abu Elzein et al. 2004). The source of the infection was not confirmed. The first reported outbreak of PPR in goats and sheep in Mongolia in 2016 resulted in spillover of infection to several species of wild ungulates, most notably Saiga antelope (Saiga tatarica mongolica). This event was associated with high mortality in the country's Saiga population, further imperiling this endangered wildlife species (Pruvot et al.
2020). While the incident did not indicate that antelope served as a reservoir or source of infection for sheep and goats, it underscored that the impact of the disease on wildlife populations must be taken into account in devising PPR control efforts. The occurrence of PPR in wild ungulates has been reviewed (Aziz-ul-Rahman et al. 2018) and guidelines for the control and prevention of PPR in wildlife populations have been developed (FAO and OIE, 2021).When populations of naive, susceptible goats are suddenly exposed to infected animals, morbidity and mortality from PPR can approach 100%. In outbreaks of PPR in endemic areas, it is young goats between 3 and 12 months of age that are most often and most seriously affected. Concurrent diseases, such as parasitism or goat pox, can increase mortality. Older animals are protected by antibody derived from previous natural exposure or by vaccination, while newborn animals up to 3 or 4 months of age have maternally derived immunity from colostrum. In regions where kidding occurs year-round, the continuous supply of susceptible young kids with waning maternal antibody facilitates maintenance of endemic PPR infection.
Seasonal peaks of disease may be observed during the heavy rains of summer or the dry cold winds of winter in some endemic countries. These increases may simply reflect peaks in susceptible young kid populations, but the pattern of disease may also be conditioned by variations in climate, geography, and differences in livestock management practices (Obi et al. 1983b; Ezeokoli et al. 1986). Pastoralism and/or transhumance that leads to high densities of susceptible animals on seasonal pastures is often associated with increases in disease. The intensification of trade in small ruminants associated with religious observance, such as Eid al Adha, may also be associated with the spread of PPiR (Sherman 2011).
Goats and sheep that recover from infection do not shed the virus, and no carrier state has been identified for PPR.
Recovered animals develop a strong immunity of at least several years' duration. Slaughterhouse surveys of apparently healthy goats in northern Nigeria indicated a seroprevalence of PPR antibody in 44% of goats tested, with goats older than 3 years of age having the highest rates of prevalence (Taylor 1979).Pathogenesis
Infected animals shed the virus in oculonasal discharges, saliva, and feces during the clinical stages of disease. The disease is transmitted by direct contact or by aerosol via the coughing and sneezing that occur. After entry into the respiratory tract, virus localizes in the tonsils and in mandibular and pharyngeal lymph nodes during an incubation period of four to six days. Viremia follows, with subsequent localization of virus in the visceral lymph nodes, spleen, bone marrow, and mucosae of the respiratory and digestive systems. Virus replication in the mucosal epithelium of the digestive tract produces mucosal erosions, resulting in stomatitis and diarrhea. Diarrhea can be severe, with many deaths resulting from dehydration and electrolyte imbalances. Virus replication in the lymphoid tissue produces a marked loss of lymphocytes. Immunity is impaired and secondary infections contribute to the severity of clinical disease and increased mortality, most notably in association with bacterial pneumonias.
Clinical Findings
Exposure to newly introduced goats or sheep during the preceding few days to weeks is a common history in explosive outbreaks of PPR. Typically, after an incubation period of four to six days, a fever of 40-41.1 °C (104-106 °F) or higher develops, and the animals become depressed, with a rough haircoat and dry muzzle. Within a few hours of the onset of fever, a serous nasal or oculonasal discharge is observed. Superficial necrosis of the lips may also be seen at this time.
The next day, affected animals become anorexic and the nasal and ocular discharges become mucopurulent. Ocular discharges are sometimes accompanied by conjunctivitis, but keratitis is not seen.
Erosive lesions may be found inside the mouth on the cheeks, lips, gums, and tongue (Figure 10.3). These lesions may be covered with necrotic tissue and inflammatory debris and there is a fetid odor to the breath. Similar erosive lesions sometimes occur on the vaginal mucosa or within the prepuce. A profuse, watery, brown diarrhea may begin one or two days after the onset of fever. Coughing and sneezing are prominent signs at this time.Fever usually persists for five to eight days; during that time, the animals grow progressively weaker and lose weight. Pregnant does may abort. The eyes, muzzle, and lips are increasingly crusted and matted and animals sneeze in attempts to clear the nares. Diarrhea persists, and marked dehydration ensues. Signs of secondary bacterial pneumonia may also develop. Although fever eventually subsides and erosive lesions in the oral cavity may heal, many animals grow steadily worse due to the persistent anorexia, severe dehydration, and complications of pneumonia. Prostration and death usually occur 7-12days after the onset of fever.
Subclinical infections are common, especially in arid regions. In these cases PPR may go unrecognized, but affected animals are likely to develop fevers and respiratory signs caused by secondary bacterial pneumonia. Movement or trade of animals with unrecognized subclinical infection is an important contributor to the spread of the disease.
Clinical Pathology and Necropsy
The hemogram demonstrates leukocytosis during the incubation period, and leukopenia during the acute phase of
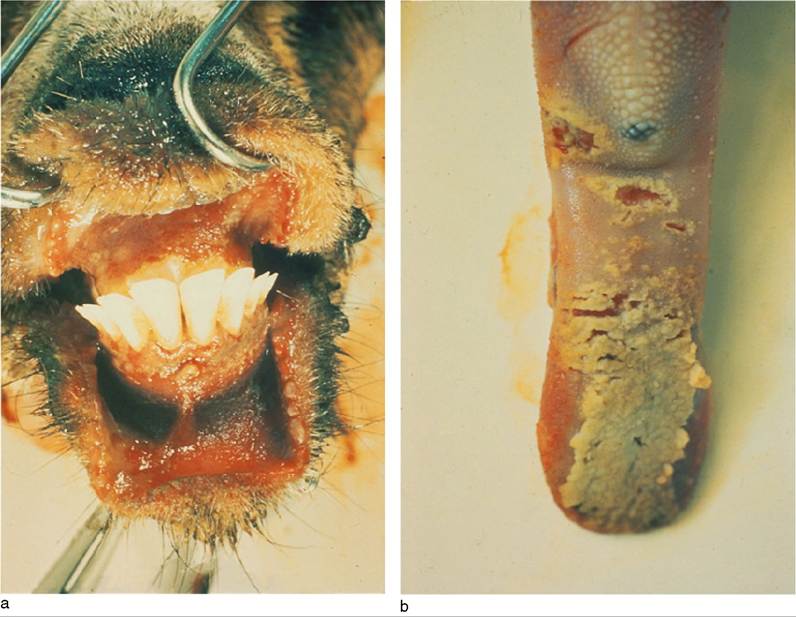
Figure 10.3 Ulceration of the gums (a) and tongue (b) of a goat with acute peste des petits ruminants. Source: From the collection of the Institute d’Elevage et de Medecine Veterinaire des Pays Tropicaux (IEMVT/CIRAD), © Dr Pierre Charles Lefevre.
clinical disease. The leukopenia is predominantly a lymphopenia. Neutrophil responses are variable depending on the development of secondary bacterial infections.
During the early febrile stage of the disease, which coincides with viremia, the virus can be isolated from the buffy coat of whole blood. Circulating antibody is not detectable during the febrile stage of the disease, but an increase in convalescent titer can be observed in recovering animals. Convalescent antibody is detectable for at least one year after recovery. Assays used for detection of antibody include virus neutralization, competitive enzyme-linked immunosorbent assay (ELISA), agar gel immunodiffusion (AGID), and counter-immunoelectrophoresis (CIEP) (OIE 2018a).
At necropsy, the carcass is emaciated, dehydrated, and soiled around the hindquarters. The eyes, muzzle, and mouth are heavily crusted. Red, raw, shallow mucosal erosions are most common on the lips, tongue, and cheeks near the commissures, and, in severe cases, on the soft and hard palate. The pharynx, esophagus, ruminal pillars, and abomasal leaves may also demonstrate mucosal erosions. Intestinal erosions are less common, but congestion is a consistent finding, with a streaky “zebra stripe” pattern of congestion or hemorrhage common in the large bowel near the ileocecal valve, cecocolic junction, and rectum. Peyer's patches may slough from the intestinal wall. Catarrhal inflammation of the upper respiratory tract with occasional petechiation and a necrotic tracheitis may be observed. The lungs may be dark red, congested, and firm. Lobular lung consolidation and airway exudate caused by secondary bacterial pneumonia are common. Lymph nodes and spleen are enlarged, congested, edematous, and possibly hemorrhagic.
Histologically, the mucosal lesions show marked epithelial necrosis with hydropic degeneration of epithelial cells at the edge of erosions. Intranuclear and intracytoplasmic inclusion bodies are common in areas of alimentary and respiratory epithelial necrosis. Syncytial cells in the oral epithelium and giant cells in lung alveoli may be observed. In the small intestine, villous atrophy and cellular casts in the crypts are seen. Lymph nodes and Peyer's patches show a dramatic depletion of lymphocytes and collapse of germinal centers. Lymphocyte necrosis is evident and macrophage numbers are increased. Multinucleated giant cells with eosinophilic intracytoplasmic inclusions are seen. Hemorrhagic necrosis occurs in the spleen.
Virus isolation should be attempted in cell culture from nasal, conjunctival, or buccal mucosal swabs in live animals or from tissues derived at post mortem, especially lymph nodes, spleen, tonsils, and lung. Preferred laboratory assays for detection of viral antigen include reverse transcriptase polymerase chain reaction (RT-PCR), realtime RT-PCR, and immunocapture ELISA, Other tests, including AGID and CIEP, are considered less reliable (OIE 2018a). An important new diagnostic development has been the validation of a lateral flow device-based test employing immunochromatographic techniques for detection of PPR antigen under field conditions (Baron et al. 2014). The availability of this penside test should greatly enhance disease surveillance efforts in support of the global PPR eradication campaign that was launched in 2015.
Diagnosis
Historically, PPR was easily confused with rinderpest, which produces a very similar spectrum of clinical signs including fever, stomatitis, and diarrhea, though respiratory signs and pneumonia, which are prominent features of PPR, were not generally associated with rinderpest. The clinical differentiation of PPR from rinderpest is no longer a practical concern, since rinderpest was declared to be globally eradicated in 2011 as a result of effective, coordinated, international control efforts (Njeumi et al. 2012).
During the early febrile stage, heartwater (cowdriosis) should be considered in the differential diagnosis. In heartwater, however, neurologic signs develop as the disease progresses. When oral lesions without diarrhea are observed, contagious ecthyma, bluetongue, and goat pox must be considered. While goats may be more severely affected by PPR than are sheep, the opposite is true in bluetongue, and goats rarely show clinical disease in endemic regions. Goat pox may present as fever followed by oculonasal discharges, but subsequent skin changes are widespread over the body.
When diarrhea predominates, salmonellosis, Nairobi sheep disease, and coccidiosis must be ruled out. In recent years, there has been an increasing overlap in the endemic ranges of PPR and Nairobi sheep disease. Coccidiosis is particularly common in the age group most often affected with PPR, but can be readily identified by examination of intestinal smears or feces.
When pneumonic signs predominate, pasteurellosis and contagious caprine pleuropneumonia need to be considered. In areas where PPR has not occurred previously, the disease is commonly misdiagnosed as pneumonic pasteurellosis. A thorough physical examination that includes inspection of the oral cavity for erosive or ulcerative lesions should help to avoid such misdiagnosis.
Treatment
In the context of the global PPR eradication effort announced in 2015, going forward animals diagnosed with PPR will likely be culled rather than treated. However, in situations where affected animals are to be treated, administration of parenteral broad-spectrum antibiotics early in the disease in affected animals can reduce the risk of likely secondary bacterial infections. Long-acting oxytetracycline, requiring an injection only every third day, is useful in this regard when repeated daily injections are impractical. The dose is 20 mg/kg given intramuscularly or subcutaneously. Oral or parenteral fluid and electrolyte therapy may reduce mortality associated with diarrhea and dehydration.
Control
Safe and effective vaccines are available for control of PPR. The most common of these is derived from a West African strain, Nigeria 75/1, but there are other attenuated vaccines produced with strains from India and elsewhere. In recent challenge studies, vaccine made with the Indian strain Sungri/96 was compared with vaccine made with the Nigerian strain 75/1, and both vaccines gave complete clinical protection against challenge with all four lineages of SRMV (Hodgson et al. 2018). PPR vaccine produced according to OIE guidelines provides protection against PPR for at least three years (OIE 2018a) and as a practical matter, under field conditions, a single vaccination is generally considered to protect animals for life.
A thermostable version of PPR vaccine has been produced (Mariner et al. 2017). There is also progress in producing so-called marker or DIVA (differentiating infected from vaccinated animals) vaccines that would permit discrimination between infected and vaccinated animals, and thereby allow countries to implement both vaccination and disease surveillance programs at the same time (Herbert et al. 2014). There are also efforts underway to produce bivalent vaccines including both PPR and capripox, as the range of both diseases overlap considerably and a combined vaccine could facilitate the logistics of large vaccination campaigns (Caufour et al. 2014). PPR, like rinderpest, is caused by a morbillivirus. The availability of long-lasting effective vaccines, the lack of a carrier state, and the apparent absence of reservoir populations for infection were all factors that supported the feasibility of the global eradication of rinderpest, a goal that was achieved in 2011. As these same conditions prevail in relation to PPR, a decision to attempt the global eradication of PPR was announced by the OIE and the FAO in 2015, and a joint PPR global control and eradication strategy was adopted with the goal of eliminating PPR by 2030 (OIE and FAO 2015). This was followed by the publication of an implementation plan for the first five years of the effort (FAO and OIE 2016).
The strategy and plan are structured around four stages. During stage one, the initial assessment stage, infected countries are expected to undertake a detailed epidemiologic and socioeconomic analysis to identify the small ruminant populations most likely involved in the maintenance and spread of PPR infection in the country, as well as the ownership, movement, and trade patterns that might foster persistence and spread of the virus. In the second, or control, stage, intensive vaccination activities are to be carried out targeting the populations of animals identified in the first phase. These vaccination efforts should be designed with the goal of achieving high levels of vaccine coverage and herd immunity in the targeted populations in order to eliminate circulation of the virus (Fournie et al. 2018; Cameron 2019). Historically, vaccination activities in some countries over many years have failed to control the disease, because low levels of vaccination were being conducted across the general small ruminant population rather than high levels of vaccination in epidemiologically targeted populations. The third stage is eradication, in which surveillance activities are intensified to ensure that vaccination has been successful, and no new outbreaks are identified. If outbreaks do occur, then stamping-out activities should be initiated as well as emergency vaccination activities if deemed necessary. Countries can enter the fourth, or post-eradication, stage once the use of vaccine has been suspended and no clinical outbreaks have been detected for 12 months. In stage 4, countries must demonstrate an additional 12 months with no clinical cases or virus circulation based on rigorous surveillance activities, and have an effective contingency plan and adequate resources to address unexpected incursions of the virus. As PPR is a transboundary disease, the activities of the OIE FAO eradication scheme are being coordinated at national, regional, and global levels.
Rinderpest
Rinderpest, or cattle plague, is best known as a devastating disease of cattle caused by a morbillivirus in the family Paramyxoviridae. Following an intensive international campaign led by the OIE and the FAO, global eradication of rinderpest was declared in 2011 (Njeumi et al. 2012). The two organizations are currently engaged in posteradication efforts to reduce and secure laboratory stocks of the rinderpest virus in high-containment storage facilities, to avoid future reemergence of the disease through accidental or intentional reintroduction of the virus.
Goats are a mildly susceptible host. Neither goats nor sheep display any ability to maintain virus transmission in the absence of large ruminant infection. Historically, the diagnosis of rinderpest in goats was complicated by the more common occurrence of PPR, a disease of similar geographic distribution, clinical presentation, and related etiology.
Reports of clinical caprine rinderpest were uncommon compared with the body of documented work in cattle. The disease was never reported as a major cause of illness of sheep and goats in Africa, even during the massive rinderpest epizootics that swept the continent in the 1890s; from then until 1955, only two more epizootics involving small ruminants were reported (Scott 1955). Over the subsequent 30 years, reports of caprine rinderpest were limited to the isolation of the virus from a kid in Uganda in 1958 (Libeau and Scott 1960) and to a putative epidemic in the eastern Sudan in 1971 that may actually have been PPR (Ali 1973; Scott 1985). A laboratory-confirmed case of rinderpest was identified in a goat in northern Pakistan in association with a severe epizootic in cattle and yaks during 1994-1995 in which more than 40 000 large ruminants died (Rossiter et al. 1998). In contrast with the lack of clinical reports, serologic surveys indicated that exposure of goats to rinderpest was common in endemic areas of Africa (Rossiter et al. 1982; Obi et al. 1984).
Unlike in Africa, outbreaks of clinical rinderpest in goats and sheep in India were reported more commonly. In the southern states of Karnataka, Andhra Pradesh, and Tamil Nadu, between 1981 and 1986 there were 984 documented outbreaks of rinderpest and 374 (38%) occurred in sheep and goats only, with mortality rates between 44.5% and 67.8% (Ramesh Babu and Rajesekhar 1988). Mortality rates as high as 80% were reported in Indian goat flocks (Mohan Kumar and Christopher 1985). It was unclear why clinical disease was more common in small ruminants in India compared with Africa. Current understanding indicates that most, but not all, of what was described as rinderpest in small ruminants was in fact PPR (Taylor et al. 2001), that if and when rinderpest did occur in small ruminants it was acquired from infected cattle with which they were in contact, and that small ruminants did not sustain infection independently (Taylor and Barrett 2007). The comparative aspects of rinderpest and PPR in small ruminants have been reviewed (Lefevre 1982; Roeder and Obi 1999).
Bluetongue
Bluetongue is an infectious, non-contagious, arthropod- borne, viral disease of ruminants. While goats are frequently infected, clinical disease is uncommon or mild in this species, even during epizootics that severely affect sheep. Direct economic losses caused by bluetongue disease in goats are minimal, although historically importation restrictions existed in bluetongue-free countries, resulting in limitations on trade of live goats, semen, and embryos. These restrictions are now becoming less stringent as scientific understanding of the disease ecology of bluetongue has improved (MacLachlan and Osburn 2006).
Etiology
Bluetongue is caused by a non-enveloped RNA orbivirus in the Reoviridae family. It has a genome of 10 segments of double-stranded RNA that produce seven structural and four non-structural proteins. For decades there were 24 different known serotypes of the bluetongue virus (BTV), distinguished by epitopes on the outer capsid protein VP2, which is encoded by the L2 gene. Individual serotypes are identified in the laboratory by serum-neutralization tests. Genetic sequencing of BTVs allows for further differentiation and analysis of strains beyond serotyping. Various serotypes may share common complement-fixing and precipitating antigens, but cross-protective immunity against heterologous serotypes is poor, creating challenges for effective control through vaccination. Virulence between serotypes varies.
Currently there are 27 serotypes identified. The newer BTV serotypes 25, 26, and 27 are referred to as small ruminant-adapted BTVs. Serotype 25 was isolated from clinically healthy goats in Switzerland in 2008. Originally referred to as Toggenburg Orbivirus (TOV), subsequent genetic characterization identified it as a distinct serotype of BTV (Hofmann et al. 2008). Experimental infection of goats with TOV produced no clinical signs in adults, but was associated with stillborn and weak kids (Chaignat et al. 2009). Serotype 26 was isolated from sheep in Kuwait (Maan et al. 2011) and serotype 27 from goats in Corsica (Zientara et al. 2014). Newer molecular technologies and metagenomic protocols continue to identify variants of existing serotypes as well as possible new serotypes, raising questions about the origin and role of these viruses in the ecobiology of BTVs (Marcacci et al. 2018).
BTV can be grown in embryonating chicken eggs, baby hamster kidney-21 cells, African green monkey kidney (Vero) cells, Aedes albopictus cells in culture, and in the KC cell line derived from Culicoides sonorensis midges (OIE 2018c). The virus is very resistant to environmental degradation. It remains infective in citrated blood held at 4 °C (39 °F) or room temperature for years. An effective disinfectant is 3% sodium hydroxide (lye) (Kohler 1989).
Epizootic hemorrhagic disease (EHD) virus is a closely related Orbivirus infecting primarily wild ruminants. It has caused disease outbreaks in whitetail deer (Odocoileus vir- ginianus) in North America. Goats challenged intravenously with EHD virus produce low levels of neutralizing antibodies, but show no evidence of viremia or clinical disease (Gibbs and Lawman 1977). Another mosquito- transmitted Orbivirus, Orungo virus, causes febrile disease in humans in Africa. There is serologic evidence of infection in goats, but clinical disease is not reported (Ezeifeka et al. 1984).
The principal insect vectors responsible for transmission of BTV are members of the genus Culicoides, variously referred to as biting midges, sandflies, no-see-ums, or gnats.
Epidemiology
Bluetongue disease was first recognized in European sheep imported into South Africa in 1870 and the disease was originally thought to be restricted to Africa. The first major epizootic confirmed outside of Africa occurred in Cyprus in 1943. The disease was identified in sheep in Texas in 1948 and the BTV-10 serotype isolated from sheep in California in 1952.
Bluetongue was first confirmed in the laboratory in Israel in 1950, but it is suspected that the disease was occurring in the Middle East since the 1920s (Shimshony 2004). Major epizootics occurred in Spain and Portugal in 1956 and 1957 and over the past 50 years there have been several incursions of bluetongue into the Mediterranean region. There have been large, notable outbreaks in Europe in recent years and the epidemiology of these European outbreaks has been reviewed (Saegerman et al. 2008). The first was in southern Europe beginning in 1998 (Gomez-Tejedor 2004), involving the Greek mainland and islands, Bulgaria, Turkey, Serbia and Montenegro, Macedonia, Croatia, and Italy. BTV-9 was mainly reported, but also serotypes 1, 4, and 16. A second wave of outbreaks occurred in 1999 in Tunisia, Algeria, and various island groups in the Mediterranean. This outbreak involved BTV-2, but serotypes 4 and 9 were also reported. Successive Mediterranean outbreaks occurred through 2004.
An outbreak due to BTV-8 began in northern and western Europe in 2006 (Elliott 2007) and continued through
2008, with no new cases in affected countries being reported after 2009. The main countries involved included Belgium, Luxembourg, the Netherlands, Germany, France, and the UK, with Denmark, Switzerland, and the Czech Republic also reporting small numbers of cases in 2007 and Norway reporting infection in four cattle herds in early
2009. A reemergence of BTV-8 in Europe began in 2015 in France and by the end of October 2020 had been reported as well in Belgium, Germany, Luxembourg, Spain, and Switzerland (Gale et al. 2020).
An outbreak caused by BTV-4 began in Greece in 2014 and by the end of the year had spread to numerous countries in southern and eastern Europe, including Albania, Croatia, Macedonia, Hungary, Montenegro, Romania, Serbia, and Turkey (Vasileiou et al. 2016). Morbidity in goats, sheep, and cattle in Greece was 2.0, 11.0, and 3.5%, respectively.
Some of these European epizootics may be facilitated by climate change as discussed further below. However, other factors may also be involved, such as wind-borne incursions of competent, infected vectors.
The global distribution of BTV reflects the distribution of competent midge vectors of the genus Culicoides. The virus is found on all continents except Antarctica. Within those continents, it is found mainly in tropical and subtropical regions. Historically the global distribution of BTV was considered to be between 40 °N latitude and 34 °S latitude, except in western North America and eastern Asia, where the northern demarcation may extend as far as 50 °N. The recent European outbreaks are noteworthy because the 1998 outbreak extended northward to 44 °N latitude in Serbia and the 2006-2008 outbreak extended above 51 °N latitude in the United Kingdom. From an epidemiologic standpoint, the geographic distribution of BTV can be categorized into three ecologic zones: enzootic, epizootic, and incursive. Enzootic zones are mainly tropical regions where BTV transmission occurs throughout the year and subclinical infection is common. Clinical disease usually occurs only when immunologically naive ruminants are introduced to the zone. Epizootic zones include temperate areas where outbreaks occur seasonally, generally in late summer when vector populations are at their highest. Incursive zones are areas that experience sporadic outbreaks when climatic conditions favor disease transmission by vectors (Gibbs and Greiner 1994).
In any given country there may be areas where infection is enzootic and areas that are free of infection. This is because local climate and geography, host populations, patterns of animal movements, and presence of competent vector species can all influence the presence of infection. For example, BTV was first isolated from a pool of trapped Culicoides in northern Australia in 1975, where the virus is enzootic (Northern Territory, Queensland, and Western Australia), but to date, except for one case in a sentinel flock maintained in the north, there are no reports of the disease in sheep, which are raised mainly in the southern portion of the country (Kirkland 2004). Similarly, in the United States, the northeast remains largely free of bluetongue, with two whitetail deer testing positive in New Jersey in 2014, while it is enzootic in much of the west and south, with seasonal outbreaks occurring in the northwest. Bluetongue in the southeastern United States has been historically limited to Florida, as the competent vector, Culicoides insignis, occurred only in the peninsula. However, recent surveys indicate that the range of C. insignis has expanded northwestward into Georgia, Alabama, Mississippi, and Louisiana (Vigil et al. 2018).
More than 1400 Culicoides spp. exist worldwide, but only 17-20 of these species have been identified as capable of BTV transmission, and the competent Culicoides spp. vectors vary regionally. Vector competency appears to be linked to BTV serotype, so that distinct regional disease ecologies or episystems exist involving particular Culicoides spp. and particular BTV serotypes (Tabachnick 2004). The current, approximate regional distribution of these vectors and serotypes is given in Table 10.4. In addition to Culicoides, the sheep ked, Melophagus ovinus, certain ticks, and biting flies have been implicated as mechanical vectors for transmission of BTV, but these are of minor importance in its dissemination. Because the virus is associated with blood cells, iatrogenic transmission by repeated use of needles when administering injections is possible.
All domestic ruminants are susceptible to BTV infection. Among sheep, European fine wool and mutton breeds are more susceptible to bluetongue than are tropical and subtropical breeds. Goats and cattle most often have subclini- cal infections and are generally considered as reservoir hosts. However, in the recent outbreaks of bluetongue in Europe, disease in cattle and to a lesser extent in goats has occurred. Even when goats are present with severely affected sheep during major epizootics, they usually have lower morbidity and mortality, exhibit milder clinical signs, or most likely remain clinically normal. The reasons for these differences in ruminant species susceptibility are not well understood.
In tropical and subtropical regions, where Culicoides are active year-round, there may be a continuous cycle of new infection in animals and the infection becomes enzootic. In temperate regions, disease is more likely to be seasonal, occurring in the summer and fall when populations of infected Culicoides have rebounded from die-offs in winter associated with cold or freezing temperatures. Vector, host, and environmental factors can influence overwintering of BTV. Culicoides do not pass BTV virus transovarially, so immature stages of the insect are not infective. Therefore, overwintering of the virus in the vector depends on the survival of at least some adult Culicoides, which can have life spans up to three months. The other major factor is the presence of virus in host animals on which Culicoides feed. The viremic period in sheep and goats is shorter than that of cattle. Most small ruminants are no longer viremic 30-40 days post infection (dpi) and cattle after 50-60 days.
In recent years, global warming appears to be altering arthropod-borne disease ecology in Europe. Increases in nighttime temperature and winter temperature along with increases in precipitation in summer and autumn have led to an increased geographic and seasonal incidence of BTV transmission by increasing the range, abundance, and seasonal activity of vectors, by increasing the proportion of the vector species that are competent, and by increasing the development rates of the virus within vectors, thereby
Table 10.4 Global distribution of bluetongue virus serotypes and the associated Culicoides vector species.
| Region | Bluetongue virus serotypes | Major Culicoides vector(s) | Other secondary or possible Culicoides vectors | Comments | References |
| North America | 2, 10, 11, 13, 17 | C. sonorensis, C. insignis (serotype 2 only) | C. variipennis | Serotype 2 restricted to Florida | Walton (2004) |
| Central America and the Caribbean | I, 2, 3, 4, 5, 6, 8, 10, II, 12, 14, 17, 18, 22, 24 | C. insignis | C. pusillus, C. furens, C. filarifer, C. trilineatus | Walton (2004) and Legisa et al. (2014) | |
| South America | 1, 2, 4, 6, 8, 10, 12, 13, 14, 16, 17, 19, 24 | C. insignis | C. pusillus | Lager (2004) and Lobato et al. (2015) | |
| Europe | 1, 2, 4, 8, 9, 16 | C. imicola | C. obsoletus, C. pulicaris, C. dewulffi | Serotypes 6 and 11 related to vaccine strains possibly used illegally | Saegerman et al. (2008) |
| Africa | 1-16, 18-20, 22, 24, 25, 26 | C. imicola | C. bolitinos and possibly others | 1, 16, 18, 19, and 24 are the predominant serotypes | Walton (2004) |
| South Asia | 1-9, 11-20, 23 | ? | C. imicola, C. oxystoma | Details on competent vector species lacking | Sreenivasulu et al. (2004) |
| Southeast Asia | 1-3, 9, 12, 14-21, 23 | C. brevitarsis | C wadai, C. actoni, C. fulvus, and possibly others | Walton (2004) | |
| Australia and | 1, 2, 3, 7, 9, 15, 16, | C. brevitarsis, | C. actoni, C. fulvus, | Serotype 2 since | Australian |
| Oceania | 20, 21, 23 | C. wadai | C. oxystoma, C. peregrinus, C. brevipalpis | 2008; serotype 5 reported in 2019 | Veterinary Emergency Plan (1996) and White et al. (2019) |
extending transmission ability to other Culicoides species. For example, Culicoides imicola, the widely distributed midge of Africa and Asia, has been the primary vector in outbreaks of bluetongue in southern Europe. The vector competence of the more temperate palearctic groups of midges, Culicoides pulicaris and Culicoides obseletus, were known from laboratory studies, but only suspected in the absence of C. imicola in outbreaks of bluetongue in Greece and southern Bulgaria in 1999 and in the Balkans in 2000-2001 (Purse et al. 2005). However, in 2006 in northwest Europe, these midges clearly established themselves as competent vectors for BTV (Elliott 2007; Saegerman et al. 2008). Culicoides obseletus was identified as the primary vector for the BTV-4 outbreak that occurred in Greece in 2014 (Vasileiou et al. 2016).
Historically, it was considered that the movement of infected livestock was a key element in the global dissemination of bluetongue and this had a significant impact on international trade in live animals, semen, and embryos, with disease-free countries prohibiting the importation of these products from countries where the disease was present. However, it is now understood that trade plays a comparatively minor role in the dissemination of bluetongue. There are two main reasons for this. The first is the recognition that viremia in ruminants has a limited duration and that persistent carriers do not occur. Some early studies, on which trade regulations were based, suggested that bovine fetuses exposed to BTV in utero become immunotolerant, persistent carriers (Luedke et al. 1977a, b). The existence of persistent carriers has since been disproven (Roeder et al. 1991; MacLachlan et al. 1994; Bonneau et al. 2002).
The second reason is the recognition that the regional occurrence of bluetongue depends on the disease ecology of the region with regard to the specific Culicoides vectors and associated BTV serotypes that occur there. There is little evidence that alien serotypes become established or persist if accidentally introduced into a region. The introduction of a new serotype to a region, such as the recent finding of BTV-8 in northern Europe, had more to do with the dissemination of competent Culicoides vectors than the movement of animals. Issues of trade policy relative to bluetongue have been reviewed (MacLachlan and Osburn 2006).
Recognition of goats as a host species for BTV was established experimentally in 1905 (Spruell 1905). The susceptibility of goats to clinical bluetongue, however, remained obscure for years, as sick goats were not observed in mixed flocks with sick sheep during outbreaks (Hardy and Price 1952).
Reports of clinical bluetongue in goats are infrequent, as the infection in goats is generally subclinical. The first reported epidemic involving goats occurred in Israel in 1950. Cattle and sheep were the principal species affected. Prevalence was less in goats and signs less severe (Komarov and Goldsmit 1951). The second reported epidemic of bluetongue affecting goats occurred in southern Spain in 1956. While losses in sheep were extreme, with more than 100 000 animals dead, cases in goats were infrequent and mild, with limited mortality (Lopez and Botija 1958). A more severe involvement of goats in a bluetongue epidemic occurred in India in 1961 (Sapre 1964). The clinical disease appeared with equal severity in sheep and goats, though variation in breed susceptibility was noted. Clinical bluetongue associated with BTV-4 occurred in goats in Portugal in December 2004, with 23 cases and 2 deaths reported to the OIE. In the epizootic in Northern Europe that began in 2006, clinical cases in goats due to BTV-8 were seen in the Netherlands (Dercksen et al. 2007; Backx et al. 2007).
The epidemiology of caprine bluetongue has been studied in the northwestern United States (Osburn et al. 1981; Stott et al. 1985). As in sheep and cattle, there is a seasonal variation in both virus isolation and serologic evidence of infection. The season of maximal infection, June through December, correlates with increases in insect vector populations. Goats are an unlikely reservoir of bluetongue infection, as no virus isolations from goats were made in California from January and June, indicating that the virus does not persist. The prevalence of infection in goats was half that recorded in cattle and sheep. All four of the major American serotypes were found in goats and multiple serotype infections occurred. However, no clinical disease was associated with infection in goats during a three-and-a- half-year period of study.
The epidemiology of caprine bluetongue has received considerable attention in Africa. In Zimbabwe, bluetongue antibody was present in 71% of goats tested, but no evidence of clinical disease has been observed (Jorgensen et al. 1989). Serologic surveys in goats in North African countries in the intertropical zone show a range of seroprevalence from 5 to 54%, with distinct differences in seroprevalence rates between different climatic regions. Low-lying and humid regions tend toward a greater prevalence rate than drier and higher regions (Lefevre and Calvez 1986). Seroprevalence in goats increases with advancing age (Obi et al. 1983a).
Pathogenesis
There are few if any studies on the pathogenesis of bluetongue specifically in the goat. Therefore, most available information is extrapolated from that on sheep or cattle. Infection results from cutaneous inoculation of the host with virus during feeding by infected insects. Initial viral replication takes place in the lymph nodes draining the feeding site and dissemination in mononuclear cells to secondary sites of replication. Viremia is usually detectable by day three and peak viremia along with fever and leukopenia usually occurs six to seven days post infection. With viremia, there is localization of the virus in vascular endothelium leading to cell damage and necrosis, with thrombosis, hemorrhage, and edema in affected organs, most notably in the tongue, mouth, esophagus, rumen, and skin, causing hyperemia, erosions, or ulcers. The virus may cross the placenta. It can also be found in semen. The clinical signs of stomatitis, glossitis, rhinitis, enteritis, and coro- nitis are attributable to the primary endothelial necrosis and loss of capillary integrity. In sheep, disseminated intravascular coagulation may occur, exacerbating the severity of clinical signs. In cattle, endothelial cell damage is minimal and the cell-associated viremia involves mainly erythrocytes and platelets (Constable et al. 2017).
Recent research on the in vitro response of ovine and bovine endothelial cells to BTV suggests that inherent species-specific differences in the production and activities of endothelial cell-derived inflammatory and vasoactive mediators contribute to the greater sensitivity of sheep to BTV-induced microvascular injury. No such comparison has been carried out yet with goat cells (DeMaula et al. 2002).
Experimental challenge studies indicate that viremia in goats can last as long as 21 days after challenge and that measurable antibody responses occur during viremia (Barzilai and Tadmor 1971; Luedke and Anakwenze 1972). Experimental challenge in goats has also confirmed that the virus can cross the placenta and infect fetuses (Coetzee et al. 2014).
Clinical Findings
In the majority of cases, bluetongue infection in goats is subclinical. When clinical illness does occur, it is often limited to mild depression, temporary loss of appetite, fever, and hyperemia of the oral and nasal mucosa.
Occasionally, more severe cases are seen. The initial signs are fever and anorexia lasting for three to four days. This is followed by hyperemia of the oral mucosa, with excoriations of the tongue, lips, and gums that become ulcerative and necrotic (Figure 10.4a). Affected goats salivate excessively. Edema of the face ensues (Figure 10.4b, c) and a watery to mucoid nasal discharge develops.
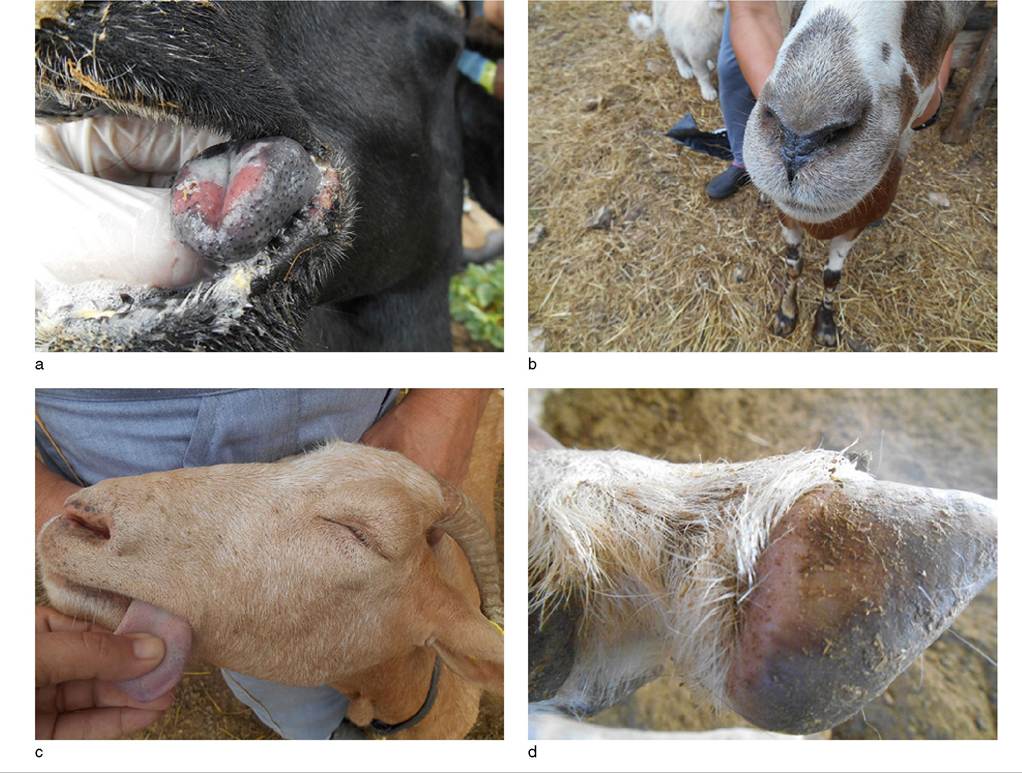
Figure 10.4 Clinical signs of bluetongue in goats observed during the BTV-4 bluetongue outbreak in Greece in 2014. (a) Swollen tongue with ulcer on dorsum. (b) Close-up of swollen muzzle. (c) Swelling of entire face, resulting in closure of eyelids and pitting edema of tongue, noticeable where the thumb is pressing. (d) Swelling and hyperemia of the heel of the foot. Reproduced by permission of Dr. Efthymios Gazgalidis.
This is sometimes followed by diarrhea. Coronitis also develops, with hyperemia and swelling around the coronet and heel (Figure 10.4d) leading to obvious lameness in one or more legs. In affected goats in India, walnut-sized skin eruptions appeared and were distributed all over the body (Sapre 1964). This presentation appears unique to goats and perhaps to the outbreak in India, because it has not been reported again elsewhere. The clinical course is 8-12 days.
Clinical cases were seen in goats in the Netherlands in August 2006, during the BTV-8 outbreak in northern Europe (Dercksen et al. 2007). The most obvious clinical signs were an acute drop in milk production and a high fever, up to 42 °C (107.6 °F). Other clinical signs were less obvious than those usually seen in sheep, but a few goats had edema of the lips and head, some nasal discharge, scabs on the nose and lips, erythema on the skin of the udder, and some small subcutaneous hemorrhages. When the field strain of BTV-8 was administered intravenously to two experimental goats, both developed viremia and antibody responses, but one remained clinically normal, while the second developed fever and became depressed, standing apart with head hanging down. It also showed dysphagia, diarrhea, and lameness. Sheep inoculated in the same experiment showed more severe clinical signs (Backx et al. 2007).
In sheep, the convalescent phase is prolonged and is characterized by emaciation and weakness secondary to active myositis. This has not yet been described in goats. Abortions have been ascribed to BTV infection in goats, but the causal role is poorly documented (East 1983).
Clinical Pathology and Necropsy
Clinicopathologic changes have not been reported in field cases of caprine bluetongue. In experimental infections, leukopenia occurs and is most pronounced 5-10 days after inoculation (Luedke and Anakwenze 1972).
Several serologic tests are available for identifying antibody to BTV. These include complement fixation (CF), AGID, and the competitive ELISA. In experimentally infected goats, antibody was first detectable at day 13 post inoculation using a blocking ELISA (Backx et al. 2007).
Virus isolation from blood and tissues is best carried out in embryonated eggs or in cell culture. However, where bluetongue is enzootic, the presence of the virus in individual animals does not establish causality when clinical signs are non-specific (Inverso et al. 1980).
The development and application of the RT-PCR has allowed for the detection of bluetongue viral nucleic acids in blood. This has revolutionized the laboratory diagnosis of bluetongue. Using this tool, antibody-positive animals can be confirmed negative for the presence of virus, allowing them to enter international trade when, in earlier times, without RT-PCR this would not have been possible (OIE 2018c).
Identification of bluetongue serogroup can be done using a number of techniques, including immunofluorescence, antigen-capture ELISA, and the immunospot test using serogroup-specific reagents. For serotyping of BTV, virus-neutralization tests are conducted in tissue culture systems using BTV serotype-specific antisera. Techniques employed include plaque reduction, plaque inhibition, microtiter neutralization, and fluorescence inhibition (OIE 2018c).
Gross necropsy lesions in goats with full-blown clinical bluetongue are similar to those observed in sheep (Sapre 1964). There is hyperemia and edema of the buccal mucosa and ulceration of the palate. Lung congestion and secondary pneumonia may be present. Severe hemorrhagic gastroenteritis may be seen in the abomasum, small and large intestines, and rectum. Pinpoint hemorrhages and congestion occur in the spleen and on the epicardium. Hydropericardium is variable. The mesenteric lymph nodes are enlarged and edematous. Hyperemia and hemorrhage may be noted on the skin of the limbs above the coronets.
Diagnosis
Definitive diagnosis of clinical bluetongue depends on isolation of the virus from animals showing typical clinical signs during seasons of insect activity. Historically, bluetongue has been recognized in goats only when clinical disease occurs simultaneously in sheep or cattle. Subclinical bluetongue in goats may be identified by changes in antibody titers in paired serum samples.
The differential diagnosis for caprine bluetongue includes foot and mouth disease, contagious ecthyma, goat pox, and photosensitization. Stomatitis and enteritis can occur in PPR, but coronitis, lameness, and edema are not prominent findings.
Treatment
There are no specific treatments for bluetongue. In sheep, prophylactic antibiotics are used to prevent secondary bacterial infections. Clinically ill animals should be kept out of direct sunlight, because sunlight appears to aggravate skin lesions. Affected animals should be fed separately so that they do not need to compete for food.
Control
Control methods are not routinely directed toward goats because the infection is generally subclinical. In general, recommendations for sheep apply. Animals should not graze in low-lying, wet areas during seasons of insect activity so as to avoid spread of infection, and paddocks and flooring of animal houses should be kept dry. Culicoides feed mostly at dusk and early evening, so animals should be removed from pastures at these times. Use of chemical insect repellants can be considered if approved for use in food animals.
In areas where seasonal occurrence of bluetongue is a known problem, vaccination is the major means of control, and vaccine should be administered prior to the onset of Culicoides feeding. Because the various serotypes offer little cross-immunity, to be effective vaccines must contain the serotypes known to be problematic in the area. In the United States, only a monovalent, modified live virus (MLV) vaccine employing serotype 10 is commercially available, although by special arrangement with the regulatory authorities, members of the California Woolgrowers Association have access to a trivalent MLV vaccine including serotypes 10, 11, and 17 (McVey and MacLachlan 2015). Multivalent vaccines against 21 serotypes are produced in South Africa. Vaccination against the full complement of serotypes requires three separate injections at three-week intervals.
There are some problems with modified-live, tissue culture virus vaccines. Passively acquired maternal antibody can impair the vaccinal response, so kids and lambs younger than 3 months of age may not be effectively vaccinated. Vaccination of ewes in the first 5-10 weeks of pregnancy can lead to fetal malformations or early embryonic death. This has not been reported in goats, but vaccination before breeding or after kidding is recommended. Vaccination should be timed to precede the insect season to promote maximum protection during high-risk periods. In addition, there are concerns of reassortment of modified live vaccine strains leading to enhanced virulence. For these reasons, there is considerable research interest in the development of new and novel bluetongue vaccines and the status of these efforts has been reviewed (Feenstra and van Rijn 2017).
Following the outbreak of bluetongue in northern Europe involving serotype 8, a number of companies in Europe produced inactivated BTV serotype 8 vaccines. Vaccination began in 2008 and proved to be highly effective in controlling the outbreak.
In the past, bluetongue-free countries had strict prohibitions on the importation of livestock, embryos, and semen, but as of 2005, the OIE revised and adopted new, less stringent international trade provisions to reflect improved understanding of the transmission of bluetongue infection, as described above. The new regulations set the infective period for bluetongue in animals at 60 days and establish protocols by which livestock, semen, and embryos can be imported into bluetongue-free countries from known positive bluetongue countries and zones through the use of quarantines and testing for antibody and virus. The availability of highly sensitive PCR tests to detect the presence or absence of virus in animals independent of their antibody status has helped to facilitate these new protocols. The current OIE provisions for bluetongue are available in the OIE Terrestrial Animal Health Code (OIE 2021).
Nairobi Sheep Disease
Nairobi sheep disease is an infectious, non-contagious, zoonotic, viral disease transmitted by ticks. Goats, sheep, and humans are affected. The disease occurs in Africa and is characterized by fever and gastroenteritis. Ganjam virus, found in India and Sri Lanka, is now considered to be the same virus as Nairobi sheep disease virus (NSDV). The relationship of the two viruses has been reviewed (Baron and Holzer 2015).
Etiology
The causative virus, NSDV, is a single-stranded, enveloped RNA virus with three genome segments (S, M, and L) of negative polarity in the genus Nairovirus of the family Bunyaviridae. There are 34 tick-borne viruses in the Nairovirus genus, classified into seven serogroups. NSDV is the prototype virus of the Nairobi sheep disease serogroup. This serogroup also includes the Dugbe virus. Earlier serologic analysis suggested that NSDV and Ganjam virus were closely related. Later sequence analysis of the S RNA genome segment and encoded proteins of both viruses indicated that they are in fact the same virus (Marczinke and Nichol 2002). The virus causing Crimean-Congo hemorrhagic fever is distinct from, but also closely related to, NSDV.
Cultivation of NSDV can be accomplished in lamb kidney, baby hamster kidney, or Vero cell culture. Intracerebral inoculation of infant mice is also used. The virus is moderately resistant to environmental degradation and can persist in blood at room temperature for as long as 45 days (Liebermann 1989).
Epidemiology
Nairobi sheep disease was first described in Africa in 1910. A major epizootic occurred when large numbers of sheep and goats were brought into Kenya in 1915 to feed troops in World War I. Nairobi sheep disease now occurs in Kenya, Uganda, Tanzania, Somalia, Ethiopia, Rwanda, Burundi, Botswana, Mozambique, and the Democratic Republic of Congo. In 1996, it was reported that NSDV was isolated from Haemaphysalis intermedia ticks collected on goats in the North Western Province of Sri Lanka (Perera et al. 1996). This same tick is responsible for transmission of the virus in India, where clinical disease has been reported in sheep and serologic evidence of infection documented in sheep and goats. The full extent of the virus as a cause of disease in small ruminants in South Asia remains largely undetermined.
Transmission of NSDV in Africa is principally by the brown ear tick, Rhipicephalus appendiculatus, in which it can be transmitted transovarially and transstadially. The distribution of the disease closely follows the geographic range of this tick and the disease is unlikely to occur in parts of Africa where R. appendiculatus has a seasonal rather than a continuous breeding cycle (Davies 1997). In Somalia, it is reported that the main vector is Rhipicephalus pulchellus, a tick that can transovarially transmit the virus (Groocock 1998). Amblyomma variegatum also transmits NSDV, but plays a minor role. A. variegatum is a less effective vector because transovarial transmission of the virus does not occur in this tick.
Infected ticks are rendered non-infectious by feeding on immune or resistant hosts. Ticks and subclinically infected sheep and goats are considered the agent reservoir. Wild ruminants and cattle are not implicated.
A seasonal occurrence of clinical disease is observed during rainy periods, when tick activity increases. Sheep and goats born in enzootic regions typically are subclinically infected and rarely show clinical disease. Clinical outbreaks are seen mainly in imported stock or stock transported from non-enzootic areas. Goats in general are less severely affected than sheep. Goat mortality may be high, but is usually less than the 50-90% mortality that occurs in sheep. Experimental inoculation of goats and sheep with blood from infected sheep resulted in lower morbidity, milder clinical signs, and lower mortality in goats (Montgomery 1917).
Human infection is acquired by tick bites. Though seroprevalence in endemic human populations is high, clinical disease is rare. In India, human disease was reported in seven laboratory personnel in association with laboratory exposure (Banerjee et al. 1979).
Pathogenesis
The pathogenesis is not completely known. The virus is introduced into goats and sheep by feeding ticks. The incubation period is 4-15 days under natural conditions and 1-3 days with experimental inoculation (Groocock 1998). Incubation is followed by viremia and localization of virus in tissues. There is an apparent predilection for epithelial cells of the gut. Bone marrow suppression also occurs. Necrosis of the mucosal epithelium of the intestines and rupture of capillaries lead to a hemorrhagic enteritis. Fluid and electrolyte loss in diarrhea is considerable and may result in death (Kimberling 1988). Recovered animals develop lifelong immunity. Kids and lambs of immune dams are probably protected by maternal antibody during the first few months of life.
Clinical Findings
Nairobi sheep disease begins with a fever of 40-41.5 °C (104-106.7 °F) that may last from three to seven days, accompanied by anorexia, pronounced depression, and a variable mucous nasal discharge that may become blood tinged. A profuse, watery diarrhea develops one to three days after the onset of fever and may progress to a bloody dysentery with straining or even some colicky signs. Depending on the severity of the disease, affected animals may die during the febrile stage before the onset of diarrhea or after one to six days of diarrhea, or they may gradually recover. Pregnant animals may abort. Swelling of the external genitalia is reported in ewes, but not in goats. Animals that recover maintain a lifelong immunity against reinfection. Reports of clinical disease in sheep or goats in India due to the virus are rare.
Clinical Pathology and Necropsy
Affected sheep and goats have a marked leukopenia during the febrile period. Serum antibody is detectable using the hemagglutination inhibition test, ELISA, complement fixation, or indirect immunofluorescent antibody test. Paired acute and convalescent serum samples should be evaluated for changes in titer. The virus may be isolated from plasma of the live animal during the febrile stage of disease. CPE in cell culture may vary with the strain of NSDV and cell type used. When CPE is absent, direct immunofluorescence staining or the indirect fluorescein antibody test confirms the presence of virus. At necropsy, virus is best isolated from spleen and mesenteric lymph nodes. A real-time RT-PCR test to detect NSDV in the blood has been described, but is not routinely available for diagnostic use. Its reported advantage is that, in comparison to cell culture, it will detect virus in blood taken after the febrile stage of disease (Baron and Holzer 2015).The AGID, complement fixation, or ELISA tests can be used to detect NSDV antigen in tissues (OIE 2018b).
Animals that die in the acute, febrile phase of the disease before onset of diarrhea may not have obvious or specific gross lesions at necropsy, and this may mistakenly lead the investigator away from a diagnosis of Nairobi sheep disease. In more advanced cases, lesions are found mainly in the gastrointestinal tract, lymph nodes, and spleen. The abomasal mucosa is hyperemic and may contain petechial hemorrhage. The gut lumen content may be bloodstained and there are numerous hemorrhages in the gut mucosa, especially in the cecum and anterior portion of the colon. These hemorrhages may have a linear or streaked appearance along the crest of mucosal folds. The spleen is considerably enlarged and engorged with blood. The mesenteric lymph nodes are also enlarged and edematous. Petechial hemorrhage may also be seen in the epicardium and endocardium and other serosal surfaces. Hyperemia of the genital tract may be noted and when fetuses are present, their tissues may show focal hemorrhages. Histologically, there is a characteristic nephritis with hyaline epithelial casts, myocardial necrosis, and hemorrhagic gastroenteritis.
Diagnosis
Nairobi sheep disease occurs only in areas where the known tick vectors occur. Disease is most likely to affect recently introduced sheep and goats. The disease should be suspected when high mortality occurs in sheep and goats with vector tick infestations, especially if the animals have been recently moved into an enzootic area or when tick populations have increased due to heavy rainfall. The definitive diagnosis is based on isolation of the virus or detection of viral antigen from clinically affected animals, or documentation of seroconversion after clinical disease.
The differential diagnosis includes PPR, Rift Valley fever, heartwater, salmonellosis, anthrax, bluetongue, and historically rinderpest, which was globally eradicated in 2011. Because fever may have subsided when diarrhea begins, coccidiosis and helminthiasis must also be considered.
Treatment
There is no specific treatment for this viral disease. Therapy should be directed at managing the effects of diarrhea and should include fluid and electrolyte therapy and adequate nutrition. Affected animals become leukopenic, so broadspectrum antibiotics are indicated to reduce the risk of secondary bacterial infections.
Control
Use of acaricides may reduce the risk of exposure for sheep and goats moving into enzootic areas, particularly following rainy seasons when tick numbers and activity increase. Vaccination of susceptible sheep and goats prior to moving them into enzootic areas has been recommended, but no vaccines are commercially available. Both an attenuated live vaccine and a killed vaccine have been developed, but there has been little demand for their use from the field (OIE 2018b).