Protozoal Diseases
Coccidiosis
Coccidiosis is the most likely cause of diarrhea in young goats between 3 weeks and 5 months of age, particularly when young goats are group housed in confinement.
Etiology
Coccidiosis is caused by protozoal parasites of the genus Eimeria. In the past, the Eimeria species infecting goats and sheep were presumed to be the same. In 1979, however, well-controlled experiments using Eimeria nina- kohlyakimovae and Eimeria christenseni demonstrated that each small ruminant host has its own host-specific Eimeria species that are not readily cross-infective (McDougald 1979). This led to a reexamination of Eimeria morphology and a renaming of species thought to be cross- infective. The nomenclature for the recognized Eimeria of goats is given in Table 10.6, with information on comparable sheep species, oocyst morphology, pathogenicity, geographic distribution, and prevalence in goats (Norton 1986; Soe and Pomroy 1992).
Historically, the Eimeria have been classified based on phenotypic characteristics such as morphology, ultrastructure, life cycles, and host specificity. However, more recent molecular phylogenetic studies of Eimeria spp. have called into question the accuracy of some of these classifications. Readers should be aware that as a result, names of Eimeria spp. affecting goats may be subject to change (Tenter et al. 2002). Furthermore, additional new species of Eimeria are being identified in goats, such as Eimeria minasensis reported from Brazil (Silva and Lima 1998) and Eimeria sundarbanensis reported from India (Bandyopadhyay 2004).
The typical life cycle of the various Eimeria spp. is as follows. Oocysts passed in the feces sporulate in the goats' environment to produce infective sporocysts containing sporozoites. These sporocysts are ingested by other goats. Ingested sporocysts release their sporozoites, which then enter host cells to form schizonts.
Schizonts undergo asexual reproduction to produce a first generation of daughter merozoites. Depending on the species of Eimeria, anywhere from a few dozen to one hundred thousand merozo- ites may be formed by each schizont. The merozoites then break from the disrupted host cell and each is capable of invading a new host cell to form a second generation of schizonts. The number of cycles of schizogony varies among Eimeria spp. The final cycle of schizogony leads to the differentiation of merozoites into male and female gametes. Male gametes (microgametes) then are released from host cells to fertilize female gametes (macrogametes) within their host cells. Zygote formation results in oocyst formation, with release of oocysts from disrupted host cells and subsequent passage in the feces.Epidemiology
Eimeria spp. have been isolated from goats on all continents. Prevalence studies throughout the world demonstrate that Eimeria oocysts are widely present in the feces of both normal and diseased goats, with a reported range of 38-100% of all goats infected (Lima 1980). Concurrent infection with multiple Eimeria spp. is the rule. For example, in a survey from Iran of 150 goats, 110 animals were infected with at least one species, but 93 of these (84.5%) were infected with multiple species (Razavi and
Table 10.6 The Eimeria species infecting goats (Capra hircus).
| Current name | Comparable Eimeria species in sheep | Morphologic characteristics of oocysts (length ? width in μm) | Prepatent period (d) | Pathogenicity | Distribution; occurrence |
| E. alijevi | E. parva | Spherical to subspherical; 16 ? 14 μm; no micropyle, no cap; yellow to yellow-green wall | 16-17 | Mild | Worldwide; very common |
| E. apsheronica | E. faurei | Ovoid; 29 ? 21 μm; distinct micropyle, no cap; brownish yellow to pink wall | 20 | Mild | Worldwide; very common |
| E. arloingi | E. bakuensis (E. ovina) | Ellipsoidal to ovoid; 27 ? 18 μm; distinct micropyle and cap | ? | Moderate to severe | Worldwide; most common |
| E. capralis | Unknown | Ellipsoidal; 29 ? 20 μm micropyle and cap; smooth wall | ? | ? | New Zealand only |
| E. caprina | None | Ellipsoidal; 32 ? 23 μm; distinct micropyle, no cap; yellow-brown outer wall, inner wall clear | ? | Moderate to severe | Known in USA, UK, Brazil; common |
| E. caprovina | Same species infects sheep | Ellipsoidal to ovoid; 30 ? 24 μm; distinct micropyle, no cap; yellowbrown inner wall, outer wall clear | 14-20 | Moderate | Known in USA, UK; uncommon |
| E. charlestoni | Unknown | Ellipsoidal; 23 ? 17 μm; indistinct micropyle; no cap | ? | ? | New Zealand only |
| E. christenseni | E. ahsata | Ovoid; 38 ? 25 μm; distinct micropyle, dome-shaped cap | 17 | Moderate to severe | Worldwide; very common |
| E. hirci | E. crandallis | Spherical to ellipsoidal; 23 ? 19 μm; flat, saucer-shaped micropylar cap | ? | Non-pathogenic | Worldwide likely; common |
| E. jolchijevi | E. granulosa | Ellipsoidal to urn-shaped; 29 ? 21 μm; distinct micropyle and cap; brownish-yellow wall | ? | ? | Known in USA, UK, Australia; uncommon |
| E. kocharli | E. intricata | Ellipsoidal; 47 ? 32 μm; distinct micropyle and light-colored cap; very thick wall | 20-27 | Mild | Known in India, Africa, Russia; uncommon |
| E. marisca | Same species may also occur in sheep | Ellipsoidal; 19 ? 13 μm; faint micropyle, shallow, domed cap; polar granule, colorless to pale yellow | 14-16 | Mild | Reported in Spain only |
| E. masseyensis | Unknown | Ellipsoidal to ovoid; 22 ? 17 μm; micropyle and cap | ? | ? | New Zealand only |
| E. minasensis | Unknown | Ellipsoidal; 35 ? 24.5 μm; micropyle and cap; two-layered wall; inner brown, outer colorless | 19-20 | ? | Reported from Brazil |
| E. ninakohlyakimovae | E. ovinoidalis | Ellipsoidal to ovoid; 23 ? 18 μm; no micropyle or cap; faint brownish- yellow wall | 15-17 | Most severe | Worldwide; very common |
| E. pallida | Same species may also occur in sheep | Ellipsoidal; 14 ? 10 μm; micropyle indistinct, no cap; yellowish green wall | ? | Non-pathogenic | Known in USA, Turkey, India, Sri Lanka; uncommon |
| Questionable status in sheep and goats | Subspherical to spherical, 21 ? 17 μm; micropyle and small cap; obvious pitting in wall and greenish tint | ? | Non-pathogenic | Reported in Germany and Zimbabwe | |
| E. sundarbanensis | Unknown | Pyriform, 28 ? 20 μm, micropyle and cap; yellowish, two-layered wall | ? | ? | Reported from India |
The species are listed alphabetically and their relative pathogenicity for goats is provided in the fifth column. Source: Based on Norton 1986; Soe and Pomroy 1992.
Hassanvand 2007). Similarly, in Zimbabwe, in a survey involving more than 1000 goats, 89.9% of adults and 94% of kids were found to be infected. All had multiple infections, with the number of species ranging from 2 to 11. Just over 75% were infected with 6-8 different Eimeria spp. (Chhabra and Pandey 1991). It is reasonable to assume that where there are goats, there are likely to be coccidia; however, it is important to discriminate between infection with coccidia and disease caused by coccidiosis.
All ages of goats may be infected with Eimeria, but numerous factors contribute to the highest incidence of clinical disease occurring in young goats between 3 weeks and 5 months of age. These include host, parasite, management, and environmental factors.
Host Factors Age-related resistance to clinical coccidiosis is reported in all ruminant species.
The resistance is immunologic in nature and is maintained by continuous exposure to coccidial infection. Immunity is relative and not absolute, because it does not eliminate infection, but effectively checks the rate of coccidial reproduction in the host intestinal tract. This is reflected in a good correlation of oocysts shed in feces with age of infected goats. There is a steady decline in oocyst numbers from 6 months through 6 years of age, followed by an increase in goats 7 years and older as immunity begins to wane in older goats (Kanyari 1988).Complete elimination of coccidial infection may lead to a loss of immunity and the development of clinical disease on reexposure to pathogenic Eimeria. The immunity that develops is specific for a given Eimeria sp. and animals of any age may develop clinical disease if exposed to a population of Eimeria spp. not previously encountered. In most management situations, goats become resistant to clinical disease at approximately 5 months of age. Resistance may be impaired, however, by stresses such as concurrent illness, lactation, transport, feed changes, nutritional deficiencies, weather changes, increased levels of exposure, or exposure to new species of Eimeria. Therefore, sporadic outbreaks of clinical coccidiosis may occur in a variety of unexpected settings, besides the predictable occurrence in confined weanlings. Breed variation may exist in resistance to coccidia. In Australia, Angora goats and feral goats are considered more susceptible to clinical coccidiosis than are dairy breeds (Howe 1984). However, a study of fecal oocyst counts in three Australian breeds showed approximately equal counts for Angoras and Anglo-Nubians, and significantly lower counts for Saanens (Kanyari 1988).
Parasite Factors Oocysts are quite resistant to environmental degradation and are even more resistant when sporulation occurs. Overwintering of sporocysts is not uncommon, and many disinfectants, including 5% formalin, do not destroy them. Sporulation depends on a combination of oxygen availability, temperature, and moisture conditions. In general, with adequate moisture and oxygen, sporulation of oocysts occurs optimally within two to five days at temperatures between 24 and 32 °C (75-90 °F) and readily at temperatures down to 12 °C (53.6 °F). Synchronous sporulation can occur among oocysts accumulating in a contaminated environment when optimal conditions prevail. This means that susceptible goats may be challenged by massive numbers of infective sporocysts under appropriate conditions of warmth and humidity.
Some of the more pathogenic Eimeria spp. such as E. ninakohlyakimovae produce thousands of merozoites per schizont during asexual reproduction inside the goat intestine. The various species may also differ in the number of cycles of schizogony involved in reproduction. The more merozoites produced, the more intestinal epithelial cells are disrupted, because every merozoite potentially can invade a target host cell. Prepatent periods also vary among goat Eimeria spp., and oocysts may be shed by kids as young as 2 weeks of age when kids are exposed to sporocysts at birth. This means that abdominal pain or diarrhea caused by coccidiosis may occur as early as 1 week of age, although this is not common.
Management and Environmental Factors Clinical coccidiosis occurs more often in intensive management situations than in extensive ones, because of the concentrating effects of confinement on both host and parasite. Dairy goat kids are commonly at risk when removed from their dams at an early age and confined to group pens indoors. Most outbreaks of clinical coccidiosis occur around the time of weaning, particularly if kids are weaned abruptly and not offered solid feed ad libitum before termination of milk feeding.
Several factors may increase exposure to infective sporocysts at this time. Feeding young goats on the ground promotes the ingestion of sporocysts. Poorly designed feeders in which goats can stand, climb, or defecate lead to contamination of feed and water sources. Watering systems that leak or overflow readily contribute to environmental moisture, which promotes sporulation. Overcrowding of pens or failure to segregate kids by age augments exposure to sporocysts and increases disease risk. When goats are kept on solid floors, the failure to maintain clean, dry bedding is a major factor in outbreaks of clinical disease. Even when floors are scraped clean, well-intentioned spraying or hosing of the pens may actually aggravate the problem because of the amount of moisture added to the atmosphere. If troughs and waterers are situated or hung so that they impede floor scraping, foci of heavy oocyst/sporocyst concentration may persist under or around them between cleanings.
Buildings that do not admit adequate sunlight also contribute to the persistence of oocysts, particularly in the winter months in temperate regions when days are short. Hot, humid weather is particularly conducive to sporocyst development and outbreaks of clinical coccidiosis are common in temperate regions during summer, especially because spring-born kids are being weaned during the hot, humid months.
Even when goats are managed extensively and the risk of coccidiosis is ostensibly less, it is important to scrutinize management practices for evidence of temporary mobbing that can predispose to outbreaks of coccidiosis. For example, Angora goats in Texas are managed extensively on range, but coccidiosis can be a major problem in weaned kids. Kids are removed from dams, placed on solid feed for the first time, and housed in large pens. Despite the spaciousness of the pens, the stressed kids flock together in one area, leading to excessive buildup of oocysts and outbreaks of coccidiosis with mortality rates as high as 15% (Craig 1986). Outbreaks in adult Angoras are also seen when inclement weather occurs after shearing, and shorn goats are confined for two weeks to prevent losses due to hypothermia. In South Africa, the incidence of clinical coc- cidiosis is aggravated by drought when great numbers of grazing animals are transferred to small, irrigated pastures or congregate to consume supplemental feed and water provided for them (van Tonder 1975).
In general, most exposures to coccidia cause subclinical infection and the acquisition of a protective though relative immunity. Most clinically affected individuals recover from clinical disease after a period of diarrhea as long as two weeks, and mortality rates do not usually exceed 10% when exposure to sporocysts has been gradual and continual before disease. When exposure to coccidial sporocysts is abrupt and intense, however, mortality rates in young goats can reach 50%. While the costs of treatment and death loss can be high, the major economic impact of coc- cidiosis in production animals may be reduced growth rates and weight gain after clinical or subclinical infection. In dairy goats, coccidiosis in weanlings can inhibit growth sufficiently that spring-born kids will not reach breeding size by the fall of the same year and must be held over an additional year. It has been reported that the oral administration of the coccidiostat decoquinate (1 mg/kg bw) to young dairy goats from eight days prior to weaning through 75 days after weaning produced higher bodyweights at 7 months of age and higher first lactation milk production at 100 and 200 days than controls (Morand-Fehr et al. 2002). In Angora goats, coccidiosis in weanlings may manifest as poor growth, poor haircoat, and increased susceptibility to pneumonia and other potentially fatal conditions.
Pathogenesis
The adverse effects of Eimeria infection in goats result from destruction of the gastrointestinal epithelium as these intracellular parasites progress through their complex life cycle in the digestive tract of the host. Multiple cycles of asexual and then sexual reproduction in the digestive epithelium result in successive waves of host cell disruption.
There are species-specific differences in the location of host cells invaded by various Eimeria spp. during their development cycles. For example, sporozoites of E. nina- kohlyakimovae, considered the most pathogenic of the goat species, enter basal epithelial cells in the crypts of Lieberkuhn in villi of the small intestine. Many thousands of merozoites are produced in each schizont. These mero- zoites then invade epithelial cells in the large intestine. The subsequent gametogonous stages invade the ileum, cecum, and large intestine, where oocysts are produced. Because multiple infections are common in goats, disruption of gastrointestinal epithelium can be very widespread.
Diarrhea results from disruption and inflammation of the intestinal mucosa. In massive infections, severe hemorrhage into the intestinal lumen can result in death caused by blood loss. In the more typical acute form of the disease, fluid and electrolyte loss result from a compromise of the normal resorptive potential of the intestinal epithelium, as well as leakage of plasma and lacteal constituents from the inflamed and disrupted mucosa. When extensive, these losses can lead to fatal systemic sequelae of dehydration, acidosis, and serum electrolyte derangement. Disruption of the mucosal integrity may also lead to increased susceptibility to secondary bacterial infection and resultant septicemia, thus increasing mortality rates. The ill-thrift and poor growth commonly seen in the aftermath of clinical coccidiosis are manifestations of prolonged malabsorption and maldigestion associated with permanent damage to the intestinal mucosa. In many cases, regeneration of a normal epithelial lining is not complete and evidence of mucosal scarring or villous atrophy may be observed at necropsy in poorly doing animals.
Clinical Findings
Most goats with clinical coccidiosis are between 3 weeks and 5 months of age. However, cases have been confirmed in adults as old as 7 years of age. Coccidiosis often occurs in conjunction with nematode infections, so attribution of clinical signs specifically to coccidiosis may be problematic, because signs of gastrointestinal parasitism due to nematodes can be similar (Valentine et al. 2007).
Subclinical coccidiosis should be suspected when complaints of poor growth, weight loss, or loss of fecal pellet formation are reported for young, susceptible animals in management situations conducive to the persistence and multiplication of coccidia.
Clinical coccidiosis may develop within one to two weeks of ingestion of a large dose of infective sporocysts. Peracute cases, caused by severe blood loss in the intestinal lumen, may present as sudden death before signs of diarrhea or abdominal discomfort are seen. The intestinal lumen may be filled with blood due to massive destruction of the intestinal mucosa.
In acute cases, early signs include decreased appetite, listlessness, weakness, and abdominal pain that may be manifested by crying and frequent getting up and lying down or by adopting a guarded stance (Figure 10.8). The feces may first be unpelleted, then pasty, and then a watery, yellowish-green to brown diarrhea develops. Fresh blood or melena may be seen. Tenesmus is less common than reported in cattle or sheep. The hindquarters and tail become stained and feces coated as diarrhea persists. If dehydration is severe, animals become recumbent with cold extremities and subnormal temperature. They eventually become moribund and die. In Australia, polioencepha- lomalacia is reported to be a common sequela to coccidiosis in goats (Howe 1980). That disease, producing signs of neurologic dysfunction, is discussed in Chapter 5.
Very young, highly susceptible animals may die of acute coccidiosis within one or two days of onset. Older or more resistant animals may exhibit diarrhea and weakness with resultant weight loss for as long as two weeks before spontaneously recovering. Animals that have recovered from

Figure 10.8 Clinical coccidiosis in a weanling goat. Note diarrhea-soiled tail and hindquarters, painful stance with tucked-up abdomen and rough haircoat. Source: Reproduced by permission of Dr. Jaroslaw Kaba, Faculty of Veterinary Medicine, Warsaw University of Life Sciences, Warsaw, Poland.
clinical coccidiosis may develop ill-thrift. They may be stunted, show a poor haircoat, and develop a pot-bellied appearance.
Clinical Pathology and Necropsy
The hemogram in coccidiosis may be normal, but a marked leukocytosis is possible when intestinal mucosal damage is extensive. Anemia is variable, depending on severity of disease, and may be masked by an increased PCV caused by hemoconcentration associated with dehydration. Varying degrees of hyponatremia, hypocalcemia, hypophosphatemia, and metabolic acidosis can occur. Hyperkalemia has been reported in goats with coccidiosis and this may be caused by compensatory efforts to counter acidosis.
The use of fecal flotation methods to detect oocysts, though often useful, is not a wholly satisfactory approach to diagnosis of clinical coccidiosis. When oocysts are not found in the presence of active diarrhea, it may be because the affected individuals are in an early stage of infection. Intestinal disruption with resulting diarrhea can occur during the schizogony that precedes oocyst formation. In these cases, direct fecal smear instead of fecal flotation is employed to identify merozoites. Conversely, oocysts may be found routinely in the feces of all normal goats older than 2-3 weeks of age. Therefore, finding oocysts in the feces of diarrheic goats does not confirm coccidiosis as the cause of illness. Performing oocyst counts is of limited value, unless the oocysts of pathogenic species are morphologically identified and quantified, because some of the less pathogenic Eimeria spp. are prolific egg producers (Aumont et al. 1984; Yvore et al. 1985). If oocyst counts are undertaken, a modified McMaster technique should be used, with a flotation solution made of magnesium sulfate at a density of 1.18 (Yvore and Esnault 1987).
Necropsy examination offers more reliable indications of clinical coccidiosis (Figure 10.9). Gross lesions of enteritis seen in the gut mucosa may range from a mild catarrh to overt hemorrhage or necrosis. In peracute cases, the intestinal lumen may contain fresh blood. Thickening of the intestinal wall caused by edema is not uncommon. The most consistent and characteristic lesion is the occurrence of multiple raised, white nodules measuring between 1 and 6 mm in diameter on the intestinal mucosa, which may be apparent even when the intestine is viewed from the serosal side. These nodules represent sites of active gametogony and on smears or histologic examination contain numerous macrogametes and oocysts. Lesions of enteritis and nodules may be seen in subclinical, peracute clinical, and acute clinical cases. The extent and severity of the lesions in conjunction with the history must be considered in establishing coccidiosis as the cause of death.
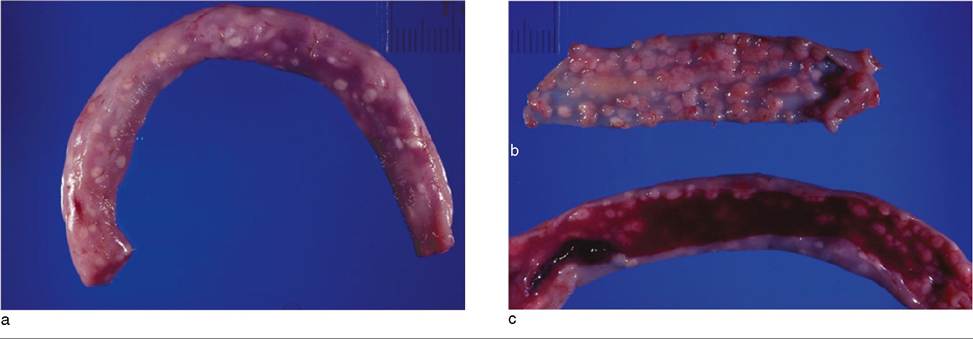
Figure 10.9 Intestinal lesions of coccidiosis in a goat. (a) Pronounced nodular lesions visible from the serosal surface. (b) Nodular lesions viewed from the mucosal surface. (c) Intestinal lumen filled with blood. This is from a weanling goat that died acutely of anemia from severe coccidiosis before the clinical onset of diarrhea. Source: Courtesy of Dr. Scott Schelling.
Lesions associated with coccidial infection have sporadically been reported in the abomasum of goats. Cysts, which appear on the mucosal surface as small whitish dots up to 1.5 mm in diameter, have been described and the condition has been referred to as globidiosis (Soliman 1960; Mehlhorn et al. 1984). Originally, Globidia was thought to be a distinct protozoal genus. It is now recognized that the protozoal structures seen histologically in the abomasa of goats and sheep are in fact large schizonts of an Eimeria species, most likely Eimeria gilruthi. Some early reports attributed diarrhea, dehydration, and death in goats to abomasal globidiosis (Mugera and Bitakaramire 1968), but the infection is also recognized to occur in normal, healthy goats (Abdurahman et al. 1987). A report of clinical disease in a sheep with coccidial abomasitis due to E. (Globidium) gilruthi suggests that the organism may indeed have pathogenic potential for goats as well (Maratea and Miller 2007).
Sporadic cases of hepato-biliary coccidiosis have also been reported in goats (Dai et al. 1991; Mahmoud et al. 1994; Schafer et al. 1995; Oruc 2007). Two types of lesions may be seen. In the biliary type, coccidial oocysts, schizonts, gametes, and gametocytes can be seen in the epithelium of the bile duct and the bile duct wall shows fibrosis. In the hepatic type, granulomata composed of oocysts and macrophages encapsulated in a fibrous capsule are present in the liver (Mahmoud et al. 1994).
Liver lesions may be severe, with large focal areas of necrosis reported (Schafer et al. 1995). Several species of Eimeria have been associated with these lesions. Clinical signs of diarrhea were reported in some of these cases, but since coccidial lesions were also present in the intestines, it was difficult to attribute the clinical disease to the liver involvement. A non-pathogenic coccidium has been identified in the gall bladder of a goat (Dubey 1986).
Diagnosis
The diagnosis of coccidiosis is presumptive, based on an appropriate signalment, history, clinical signs, and necropsy findings. In very young kids between 1 week and 1 month of age, other causes of abdominal pain and acute diarrhea must be differentiated. These include cryptosporidiosis, colibacillosis, enterotoxemia, salmonellosis, yersiniosis, viral enteritis, and dietary diarrhea. Abdominal pain alone would suggest abomasal bloat, mesenteric torsion, or other intestinal accidents if diarrhea does not develop. In cases of peracute death due to coccidiosis, the differential diagnosis includes enterotoxemia, bacterial septicemias, and plant and chemical toxins.
In weanlings 2-5 months of age, particularly those with access to pasture, kid yards, or poorly drained dry lots, helminthiasis is the major differential diagnosis for diarrhea. Gastrointestinal nematodiasis and paramphistomiasis should be considered. In fact, multiple parasitic infestations are common and it may be difficult to attribute diarrhea or ill-thrift to coccidiosis alone. Other causes of diarrhea in this age group include simple indigestion and acute carbohydrate engorgement due to sudden feed changes or excessive grain feeding, as well as salmonellosis and enterotoxemia.
In subclinical coccidiosis, or after severe clinical coccidi- osis, ill-thrift or poor growth may be the primary clinical complaint in growing kids. Selenium deficiency, copper deficiency, or cobalt deficiency must be considered in these cases, particularly where soil deficiencies of these minerals are known to exist. Chronic helminthiasis must also be ruled out.
Treatment
Supportive care is the principal therapeutic intervention in active cases of coccidiosis. Diarrheic goats should be removed from the group and given oral or parenteral balanced electrolyte solutions, depending on the degree of dehydration. In preweaned kids, milk should be fed only in small amounts, because disruption of the intestinal mucosa produces maldigestion and may promote osmotic diarrhea from undigested lactose. Adding commercially available lactase enzyme (e.g., Lactaid®, McNeil Nutritionals, Ft. Washington, PA, USA) to the milk may also improve tolerance. Weaned kids should be offered good-quality grass hay and brought back on full feed gradually. Severely anemic kids with acute intestinal blood loss may require blood transfusions. Broad-spectrum antibiotics are indicated in severe cases to prevent bacterial septicemia secondary to disruption of the intestinal mucosal barrier. Probiotics have been recommended to help reestablish the normal gut flora (Bath et al. 2005).
The use of anticoccidial drugs in active clinical cases may have limited value. Most anticoccidial drugs are coc- cidiostats that inhibit but do not eliminate coccidial reproduction. In fact, shedding may be higher several weeks after treatment than it was before. These drugs usually act on early stages of the reproductive cycle. Animals already showing diarrhea have often progressed beyond the stages of infection where coccidiostats may prove beneficial. Nevertheless, goats are commonly infected with multiple species of Eimeria and some of these may still be in the early stages of development. In these cases, the duration of clinical disease may be shortened by treatment.
The main goal in administering coccidiostats is to reduce the number of additional cases developing in a group of animals at risk, rather than curing existing cases. Drugs reported for use in the treatment of caprine coccidiosis include sulfonamides, nitrofurazone, ionophores, ampro- lium, toltrazuril, and diclazuril. Dosages and treatment regimens reported specifically for goats are summarized in Table 10.7.
The sulfonamides and nitrofurazone may offer the added benefit of control of secondary bacterial infections. Because goats with clinical coccidiosis are often dehydrated, the sulfonamides must be used with care, because nephrotoxicity can result from poor renal perfusion and decreased urine output. Sulfonamides are the oldest class of drugs used as coccidiostats and it is generally considered that resistance to sulfonamides is widespread among Eimeria spp. However, efficacy continues to be reported in some goats treated with sulfonamides. Sulfa drugs are available for oral and parenteral use. A variety of oral preparations can be given as boluses or added to feed or water. Nitrofurazones can produce neurotoxicity when overdosed, as may occur if the medicine is not properly mixed into milk or solid feeds. This drug is rarely used any longer in coccidiosis control. Note that in 2002, as a public health measure, all nitrofurans, including nitrofurazone, were banned in the United States for use in food-producing animals, because the nitrofurans have been identified as potential carcinogens.
Amprolium is a thiamine antagonist and works as a coc- cidiostat by blocking utilization of thiamine by coccidia. If amprolium is used for too long or at excessive doses, there is the potential for producing polioencephalomalacia in goats, though this should not discourage its proper usage. Polioencephalomalacia of ruminants may result from thiamine deficiency, as discussed further in Chapter 5. Amprolium is for oral use and is available as a liquid for addition to milk, milk replacer, or water, and in solid forms for addition to feeds.
The ionophores include monensin, lasalocid, and salino- mycin; all have been used in goats. They are used mainly in solid feed in weaned animals. While monensin is used widely in poultry, cattle, and sheep, it is known to be toxic in horses and is potentially toxic in ruminants if overdosed. The lethal dosage LD50 in sheep is reported to be 12 mg/kg; in cattle, 22-80 mg/kg; and in goats, 26.4 mg/kg. Muscle weakness and myoglobinuria are the clinical signs of toxicity in sheep (Langston et al. 1985). In a toxicity study, goats given monensin at a dose of 13.5 mg/kg daily for five consecutive days via gastric gavage showed effusions in body cavities, vacuolar degeneration and coagulative necrosis in cardiac and skeletal muscles, and renal tubular necrosis at necropsy (Deljou et al. 2014). The cardiotoxicity of ionophores is discussed further in Chapter 8.
Another class of coccidiocidal drugs known as the symmetrical triazinons emerged in the 1980s. They can disrupt all intracellular developmental stages of Eimeria and therefore can be more effective in the treatment of clinical coc- cidiosis than are the coccidiostats, while also being useful for prophylaxis. Two drugs in this class are used in ruminants in other countries, toltrazuril and diclazuril, but neither is specifically approved for use in ruminants in the United States. Diclazuril and a third drug in this class, ponazuril, are approved for use in horses in the United States, but extralabel use in goats is not permitted.
There are several reports on the use of toltrazuril in goats. In the earliest report, a single oral dose of 20 mg/kg produced a rapid, significant reduction in oocyst shedding that remained low for two to three weeks, suggesting that all developmental stages of the coccidia present were killed (Anonymous 1988a). In a later study, toltrazuril was administered orally at a dose of 25 mg/kg for two consecutive days to 7-week-old kids with clinical coccidiosis. Oocyst shedding was virtually eliminated and all treated kids became clinically normal (Ocal et al. 2007). In another
Table 10.7 Coccidiostats used in the treatment or prevention of caprine coccidiosis.
| Drug used | Treatment regimen | Prophylaxis regimen | Comments | References |
| Sulfonamides | ||||
| Sulfadimethoxine | 75 mg/kg bw orally for 4-5 days | Yvore (1984) | ||
| 250 mg/kg bw orally once using a sustained-release bolus | Releases 50 mg/kg bw per day for 5 days | Yvore et al. (1986) | ||
| Sulfadimidine | 135 mg/kg bw orally for 4-5 days | 55 g/t of feed for at least 15days | Yvore (1984) and Constable et al. (2017) | |
| Sulfaguanidine | 280 mg/kg bw orally for 4 days | Vujic and Ilic (1985) | ||
| Sulfaguanidine (or Sulfaquinoxaline) followed by Sulfathiazole (or Sulfadimethoxine) | 1.3 g/kg bw orally for 4days 1.1-2.2 g/kg bw orally for 4 days | Poor absorption from digestive tract | Guss (1977) | |
| Sulfamethazine | 50 g/t of feed | Shelton et al. (1982) | ||
| Antibiotics | ||||
| Nitrofurazone | 10-20 mg/kg bw orally for 5-7 days | No longer permitted for use in food producing animals in the USA | Tarlatzis et al. (1955, 1957) | |
| Amprolium | 10-20 mg/kg bw orally for 3-5 days | Yvore (1984) | ||
| 50 mg/kg bw orally for 5 days | 25-50 mg kg bw orally in feed or water continuously from 2 weeks to several months of age | Swarup et al. (1982) Smith (1980) | ||
| Ionophores | ||||
| Monensin | 15-20 g/t of feed | Shelton et al. (1982) and Foreyt (1990) | ||
| Lasalocid | 20-30 g/t of feed (1.1 2.2 mg/kg bw/day) continuously | Williams (1982) | ||
| Salinomycin | 100 ppm in concentrate fed for three weeks after weaning | Yvore (1984) | ||
| Quinolones Decoquinate | 0.5-1 mg/kg bw orally in feed; may be fed continuously | Foreyt et al. (1986) | ||
| Triazinons | ||||
| Toltrazuril | 20 mg/kg bw orally once | 20 mg/kg bw orally once every 3-4 weeks | McKenna (1988) | |
| Diclazuril | 1 mg/kg bw orally once | 1 mg/kg bw orally once every 3-4 weeks | Recommended for use in goats but no dose given. Lamb dose provided here | Harwood (2004) |
| Ponazuril | 10 mg/kg bw orally once | Gibbons et al. (2016) | ||
| Clopidol and methylbenzoquate | 12 mg/kg bw clopidol and 1 mg/kg bw methylbenzoquate/day for 5weeks | Polack et al. (1987) | ||
| Dapsone | 80 mg/kg bw orally for 4 days | Devillard (1981) | ||
| 3,5, Dinitro-O-toluamide | 100 mg/kg orally for 5 days | Dash and Misra (1988) |
bw, bodyweight; ppm, parts per million.
study, the drug was given to 4-5-week-old kids at a lower dose of 10 mg/kg bw for two consecutive days (Slosarkova et al. 1998). Though oocyst counts in feces were reduced, considerable numbers of oocysts continued to be shed over the following three weeks and by five weeks were at higher levels than the pretreatment counts. Therefore, the higher doses of 20-25 mg/kg appear to be more effective in kids and a single oral administration is sufficient to be effective (Arunvipas et al. 2013; Nunes et al. 2015).
Diclazuril has been recommended for prophylactic use in goats. It is available in some countries as an oral suspension and is given as two doses two weeks apart (Harwood 2004). In lambs, the therapeutic and prophylactic doses of diclazuril are the same, 1 mg/kg bw, given at 4-6 weeks of age, with a second dose administered three weeks later for prophylactic use where there is high infection pressure.
Ponazuril has been evaluated as a treatment for coccidi- osis in goat kids in comparison to amprolium. Given one time at a dose of 10 mg/kg orally, ponazuril reduced oocyst counts in feces to the same extent as amprolium given daily for five days at an oral dose of 50 mg/kg per day (Gibbons et al. 2016).
In addition to the use of coccidiostats, implementing management changes is important in controlling outbreaks of coccidiosis. Attempts should be made to reduce exposure to infective sporocysts by removing contaminated bedding and feed, reducing the stocking rate, and moving animals to a new, uncontaminated environment.
Control
Under management conditions where outbreaks of coc- cidiosis can be reliably anticipated, measures should be taken to control coccidiosis before clinical disease is observed. Proper control is a balancing act. The goals are to reduce oocyst numbers in the environment sufficiently to avoid massive challenge to susceptible animals, while allowing sufficient exposure so that immunity and resistance can properly develop. This is accomplished by combining sound management and hygiene practices with the use of coccidiostats. In production systems where group housing of weanlings is unavoidable, control of coccidiosis without the use of coccidiostats is rarely achieved, even when management is excellent. This is especially problematic in US organic systems, where coccidiostats are not permitted.
In attempting to maximize the immunologic resistance of the host, it is important to consider the mode of action of the various coccidiostats. The host immune response appears to be most stimulated by active schizogony, while most coccidiostats inhibit reproduction earlier in the infective cycle.
Sulfonamides, amprolium, and ionophores have been used as prophylactics and as therapeutics in goat-rearing programs, with mixed success. The sulfonamides are folic acid antagonists and are most inhibitory during the second schizogony. This late action allows for the development of host immunity. However, as already mentioned, resistance to sulfas may be widespread. The ionophores primarily inhibit the early, asexual stages of coccidial development. They offer an added benefit of improved weight gain above and beyond their action as coccidiostats (Shelton et al. 1982). Unfortunately, outbreaks of clinical coccidiosis have been observed in young goats being fed lasalocid in their ration. This has been attributed to uneven consumption by individual goats, possible resistance to the drug, and failure to maintain good hygiene, in addition to feeding coccidiostats. Monensin was approved for use in non-lactating goats in the United States in 1989 as a feed additive at 20g∕t.
Decoquinate, a quinolone not used for treatment, has been found to be effective in preventing caprine coccidio- sis, and it is very safe, with no LD50. It works early in the Eimeria life cycle by inhibiting sporozoite development. Dose ranges from 0.3 to 4 mg/kg bw have been evaluated. All doses prevented clinical coccidiosis and allowed equal rates of weight gain. Doses at the higher end, however, resulted in a more rapid and pronounced decrease in the number of oocysts shed (Foreyt et al. 1986). In 2002, deco- quinate was approved for use in non-lactating goats in the United States as an additive to milk replacers or feeds, to be administered at a dose of 0.5 mg/kg bw for a minimum of 28 days to control coccidiosis due to E. ninakohlyakimovae and E. christenseni (Ecfr.gov 2022).
Coccidiostats are most often used in goats between the ages of 1 and 4 months, usually in conjunction with grain feeding. However, in some intensive management situations, it may be necessary to begin use in the milk or milk replacer of preweaned kids. As discussed above in the treatment section, in countries where use is allowed, oral toltrazuril or diclazuril can also be highly effective for prevention of clinical coccidiosis in young kids if given at 3-4week intervals through weaning.
Good hygiene and thoughtful management are essential to effective coccidiosis control. Weaning should be as unstιress- ful as possible. To minimize weaning shock, kids should have access to grain well in advance of weaning. Whenever possible, dairy kids should be raised away from adults and housed individually or in small groups, segregated by age, and provided feed and water in devices that minimize contamination with feces. They should never be fed on the ground, and waterers must not leak or be liable to spillage.
Housing that allows shelter with ready access to sunlight is preferred. A successful system of weaned kid management using movable hutches is shown in Figure 10.10. No

Figure 10.10 Movable hutches used in coccidiosis control. Fencing and hutches are moved every several weeks to minimize weanling exposure to oocyst buildup on the ground. Source: Courtesy of Vincent and Christine Maefsky, Poplar Hill Dairy Goat Farm, Scandia, Minnesota.
matter what type of housing is used, ground, floors, and bedding should not be allowed to become excessively wet. Adequate ground drainage, frequent floor scraping, and regular provision of clean bedding are necessary. Scraping floors and allowing them to dry is probably more effective in coccidia control than scraping followed by scrubbing or hosing, because disinfectants are generally not effective, and excess moisture promotes sporulation. If disinfectants are used, products containing ammonia, particularly quaternary ammonium compounds, are the most effective.
In general, when proper control methods are implemented and goats are not exposed to new species of Eimeria not previously experienced, immunity to reinfection is usually achieved by 5 months of age if the animals have good nutrition. Coccidiostat feeding may be terminated and, if possible, kids should be moved to uncontaminated housing at this time.
No vaccines are currently available for control of coccidi- osis in ruminant animals, though live, multivalent, attenuated coccidial oocyst vaccines are commercially available for poultry. Preliminary studies have demonstrated that oral administration of X-irradiated, attenuated E. nina- kohlyakimovae oocysts to goats can effectively immunize them against homologous infections (Ruiz et al. 2014). There has been active research into the development of subunit vaccines for protozoa of veterinary importance, including the Eimeria, and these efforts have been reviewed (Jenkins 2001).
Cryptosporidiosis
First reported as a cause of fatal enteritis in goats in 1981, cryptosporidiosis has emerged as a major cause of diarrhea in kids younger than 1 month of age, particularly under intensive management conditions. Cryptosporidiosis is a zoonotic disease that causes vomiting and diarrhea, especially in children. The occurrence is often associated with direct contact with livestock on farms or at petting zoos, or the use of recreational water sources. It is also an important and serious infection of patients with acquired immunodeficiency syndrome (AIDS) or other conditions involving immunosuppression. The disease in goats has been reviewed (Paul et al. 2014).
Etiology
Cryptosporidiosis is caused by Cryptosporidium spp., which are small, protozoal parasites in the family Cryptosporidiidae in the phylum Apicomplexa. Though long considered to be members of the class Coccidea, they are no longer considered to be coccidian parasites (Xiao and Cama 2007). The taxonomy of the Cryptosporidium genus is in flux as new molecular criteria become available to augment or replace the more traditional morphologic criteria of classification. Currently, close to 40 Cryptosporidium species have been identified that infect a wide range of vertebrate hosts, including humans; all the mammalian domestic livestock species; birds, including domestic poultry; reptiles; and fish. These species tend to be, but are not exclusively, host specific. Cryptosporidium parvum infects domestic ruminants, including goats, sheep, and cattle as well as humans, and is the major focus of this discussion. In addition to C. parvum, other species of Cryptosporidium identified in goats from different areas of the world include Cryptosporidium andersoni, Cryptosporidium hominis, Cryptosporidium ubiquitum, and Cryptosporidium xiaoi (Hatam-Nahavandi et al. 2019). It is known that cryptosporidial oocysts are very resistant to environmental degradation, and many common disinfectants are reported to be ineffective in destroying them. Oocysts remained infective after 18 hours of contact with iodophors, cresylic acid, sodium hypochlorite, benzalkonium chloride, and sodium hydroxide. Only 5% ammonia and 10% formol saline destroyed the infectivity of oocysts (Campbell et al. 1982). Ammonium hydroxide, hydrogen peroxide, and chlorine dioxide are now also recognized to be effective disinfectants.
Epidemiology
Cryptosporidiosis occurs worldwide. In goats, kids from 2 days to 3 weeks old are mainly affected, but older animals may also be affected. In outbreaks, especially in intensively managed herds, kid morbidity can approach 100% and mortality up to 40% has been reported.
The first reported field case of diarrhea attributed to cryptosporidiosis in a goat involved a 4-week-old kid in Australia in 1978 (Mason et al. 1981). This was followed by a second Australian report in 1982 in which 21 of 29 grazing goat kids developed diarrhea and 3 died. All kids examined were shedding Cryptosporidial oocysts and tests for enteric viruses and K99+ Escherichia coli were negative (Tzipori et al. 1982).
In Hungarian studies on the causes of diarrhea in kids younger than 1 month of age in large commercial dairy goat herds, cryptosporidia were isolated more frequently than rotavirus, coronavirus, adenovirus, or enterotoxigenic E. coli (ETEC). In these studies, cryptosporidia were not identified in the feces of any normal goats, while all of the other agents mentioned were identifiable one or more times in non-diarrheic kids (Nagy et al. 1983, 1984). In a French survey, 28 of 48 diarrheic kids (58%) on nine different premises had cryptosporidia detectable in feces or intestinal scrapings (Polack et al. 1983). A more extensive kid survey on 24 intensively managed dairy goat farms in France demonstrated an overall mortality rate from diarrhea of 10.3%, mostly during the first two weeks of life. Cryptosporidia were identified in feces of virtually all fatal diarrhea cases, but not in any non-diarrheic animals, save those recovering from diarrhea or those that subsequently developed diarrhea (Yvore et al. 1984). In contrast, cryptosporidia have been reported from feces of healthy kids in Tanzania (Matovelo et al. 1984) and Nigeria (Ayeni et al. 1985).
In diagnostic laboratory accessions seen from 1981 to 1985 in New Zealand, cryptosporidia were identified more frequently than any other single infectious agent, save Eimeria spp., in diarrheic kids younger than 1 month of age (Vickers 1986). Cryptosporidia also have been reported in association with diarrhea in a 6-month-old goat in the Netherlands, 4-25-day-old kids in Italy, and a 7-day-old kid in conjunction with C. perfringens in the United States (Ducatelle et al. 1983; Gialletti et al. 1986; Card et al. 1987). The author (DMS) has confirmed cryptosporidiosis as a significant cause of diarrhea in young, intensively managed kids in a large goat herd in New York. Major outbreaks of cryptosporidiosis affecting kids by the hundreds have been reported in herds from Oman (Johnson et al. 1999) and Turkey (Sevinc et al. 2005). In the Turkish incident, subclinically infected adult goats, insufficient colostrum intake in kids, and contamination of the kidding area were considered to contribute to the dramatic evolution of the outbreak.
Transmission is by the fecal-oral route from oocysts shed in the feces, and infections can spread quickly through group-housed kids once introduced into the group. It has been shown that goats may be infected with cryptosporidia isolated from calves and humans (Contrepois et al. 1984; Nagy et al. 1984). Colostrum-deprived kids develop more severe disease and a higher rate of mortality than do colostrum-fed kids (Naciri et al. 1984). It is not yet clear if this is directly associated with Cryptosporidium-specific immunoglobulins or with other non-specific nutritional or immune factors available from colostrum. The severity of disease does not appear to be affected by the state of the intestinal flora (Contrepois et al. 1984). While cryptosporidia are found infrequently in non-diarrheic kids during field investigations, experimental challenge can result in oocyst shedding in diarrhea-free kids, indicating that subclinical infections may occur (Nagy et al. 1984).
Pathogenesis
Cryptosporidium spp. are obligate, intracellular parasites. The only stage that occurs outside the host is the oocyst stage. The life cycle is similar in outline to the Eimeriidae, with some notable differences. Transmission is by the fecal-oral route. Susceptible hosts ingest infective, sporu- lated oocysts, which excyst and release sporozoites that invade intestinal enterocytes and undergo rapid asexual multiplication or schizogony. Meronts produced by schizogony release merozoites, which invade new enterocytes. Sexual multiplication or gametogony also occurs in entero- cytes, producing zygotes. Zygotes may form either thinwalled sporulated oocysts, which rupture within the small intestine to release sporozoites that can further invade new host enterocytes (autoinfection), or a thick-walled sporu- lated oocyst, which is passed in the feces as a source of infection to other animals.
Initially, these parasites of the gastrointestinal tract were considered to be extracellular, observed in close association with the brush border of the intestinal epithelial cells. Electron microscopic studies have confirmed that they are in fact intracellular, but extracytoplasmic, residing underneath the brush border of intestinal epithelial cells in a parasitophorous vacuole created from the host cell microvillous and plasma membranes.
The pathogenic effects of cryptosporidial infection are most likely due to the impairment of enterocyte function and integrity resulting from the rapid, cyclic reproduction of cryptosporidia that occurs within the brush border of enterocytes. In goats, infection leads to increased senescence of epithelial cells and villous atrophy in the ileum. The severity of these effects is related to the degree of infection (Matovelo et al. 1984). This can lead to both malabsorption and maldigestion, resulting in the clinical diarrhea observed. Prolonged weight loss may be associated with malabsorption. Steatorrhea caused by maldigestion has been documented in experimentally infected lambs (Contrepois et al. 1984). That diarrhea has been observed in kids as young as 3-4 days of age reflects the short life cycle of cryptosporidia. The prepatent period has been reported to be two to seven days in calves and two to five
days in lambs (Constable et al. 2017). Age-related immunity probably occurs, since clinical disease is far less common in older animals, but this immunity is not complete, as oocysts are found in the feces of healthy adult goats (Castro-Hermida et al. 2007a). Such goats can serve as a source of infection for kids. In ewes, it has been reported that there is a periparturient increase in the excretion of oocysts.
Clinical Findings
The most common presentation of cryptosporidiosis is an acute, white to yellow, watery diarrhea in kids younger than 2 weeks of age. The diarrhea may last from several days to two weeks, and may be mild to severe, presumably depending on the magnitude of the initial exposure to oocysts. Diarrhea may be persistent or recurrent. In addition to diarrhea, kids may show depression, inappetence, and a rough haircoat. Sequelae of dehydration, electrolyte imbalance, acidosis, and death may occur, depending on the severity of diarrhea and the time of intervention. Spontaneous recovery can also occur.
Explosive outbreaks of diarrhea with mortality rates of 10-20% have been observed in intensively managed dairy goat kids housed in groups indoors. Some of these outbreaks have begun toward the end of the kidding season, suggesting that environmental buildup of oocysts is contributory (Delafosse et al. 2006).
Two other syndromes of cryptosporidiosis have been reported from France (Polack and Perrin 1987). Progressive emaciation without diarrhea has been observed starting in 1-week-old kids, and diarrhea has been noted beginning in 6-week-old goats.
An atypical presentation of caprine cryptosporidiosis was reported from Oman (Johnson et al. 1999). Distinguishing features included clinical disease in adult goats as well as kids, and the limitation of cryptosporidiosis to goats, even though sheep, cows, and buffalo were in close proximity on the farm. No other enteric pathogens were identified during the outbreak. Morbidity approached 100% in goats under 6 months of age and mortality was high, with 37.8% of goats up to 4 weeks of age and 34.4% of goats between 5 and 8 weeks of age dying.
Clinical Pathology and Necropsy
Current evidence suggests that cryptosporidial oocysts are not commonly shed in large numbers in the feces of healthy goats. Detection of oocysts in feces of diarrheic goats therefore is useful in supporting a diagnosis of cryptosporidiosis. Direct fecal smears have been routinely used for examination because they are quick and easy to prepare. Oocysts, however, may be sparse, difficult to discover in fecal debris, or hard to distinguish from yeasts. Fecal flotation and sedimentation methods are preferable. Centrifugal flotation techniques including the use of potassium dichromate/ saturated sucrose (specific gravity [sp. gr.] ≥ 1.10), sucrose/ phenol (sp. gr. = 1.27), and potassium iodomercurate (sp. gr. = 1.44) solutions have been reported. Yeasts do not float in the potassium dichromate/saturated sucrose solution (Willson and Acres 1982).
Concentrated oocysts can be viewed stained or unstained using direct light- or phase-contrast microscopy. The sample for examination should be taken from the meniscal surface using a wire loop, put on a slide, and covered with a coverslip. It is advisable to use high-power magnification, search the edges of the coverslip, and focus just below the coverslip, because oocysts float up to its underside. A semi- quantitative flotation technique for counting oocysts in feces has been reported for use in calves (Anderson 1981) and applied to goats (Contrepois et al. 1984). Sedimentation using formalin-ethyl acetate solution with centrifugation at 1700 ? g or more also effectively concentrates oocysts (Kirkpatrick and Farrell 1984).
Numerous staining techniques have been used to assist in identifying cryptosporidia oocysts in feces. Currently, acid-fast staining with modified Ziehl-Neelsen stains is generally favored. Oocysts stain pink to red, with interior blue granules against a blue-green counterstained background. Auramine fluorescent stains or fluorescein isothiocyanate-labeled monoclonal antibodies are also being used. With these stains, oocysts display a yellow green fluorescence against a black background. Antigenbased ELISA tests and PCR techniques are also available for detection of C. parvum oocysts in feces (Wright and Coop 2007). Immunoassays are available too, but are more appropriate for surveillance studies than for case diagnosis, because clinical disease generally precedes the immune response.
Rapid field diagnostic tests also have become available. Based on lateral flow immunochromatographic techniques, antigens of C. parvum in feces can be detected on monoclonal antibody-impregnated test strips through formation of a positive, colored test line after five minutes of incubation.
Gross lesions are non-specific and unremarkable, and may be limited to a mild to moderate enteritis most commonly located in the ileum. Smears should be made of the ileal mucosa for microscopic examination to detect oocysts. Histologically, sections of intestine reveal numerous small, round basophilic bodies of various sizes along the mucosal surface. These are the different endogenous stages of developing cryptosporidia in the brush border of the entero- cytes. Free oocysts may also be seen in the intestinal lumen. Infection is accompanied by marked villous atrophy and villous fusion in the ileum, and possibly the jejunum and large intestine. For accurate diagnosis, it is essential to fix fresh tissues in formalin immediately after death, because autolysis of developing cryptosporidia and associated enterocytes is rapid. Oocysts in feces are hardier, although freezing destroys them. If necessary, they may be preserved as long as 120 days by mixing one part feces with two parts of 2.5% potassium dichromate solution.
Diagnosis
A presumptive diagnosis of cryptosporidiosis is based on identifying oocysts in feces or various endogenous stages of the organism in intestinal smears or histologic sections. Since cryptosporidia may be present in association with other etiologic agents capable of producing diarrhea, laboratory evaluation for the various other bacterial, viral, and protozoal causes of kid diarrhea is indicated before a definitive diagnosis can be made. These are discussed later in the chapter in the section on the neonatal diarrhea complex, and a list of likely etiologic agents is provided in Table 10.3. Careful history taking is necessary to rule out dietary causes of diarrhea.
Treatment
Currently, treatment of cryptosporidiosis is limited to supportive care, particularly oral or parenteral fluid therapy, based on the severity of fluid and electrolyte loss. Nutritional management is an important consideration, because maldigestion may be present. Diarrhea in cryptosporidiosis can last as long as two weeks. It is not possible to withhold milk for that period, but regular milk feeding may result in aggravation of the diarrhea due to the osmotic influence of undigested lactose. Therefore, it is recommended to reduce the volume of milk fed per feeding while increasing the frequency of feeding. The use of commercial, lactose-free milk products or the addition of commercially available lactase enzyme (e.g., Lactaid) to goat milk before feeding to kids can be helpful. Lactose intolerance has been documented in a young goat in association with clinical cryptosporidiosis and feeding of lactase-treated goat milk was reported to aid in recovery of the affected kid (Weese et al. 2000). Parenteral nutrition, though costly, could be considered in pet goats or other valuable individuals.
Reportedly, more than 50 anticoccidial, antibacterial, and antiparasitic drugs have been tested for therapeutic efficacy against cryptosporidiosis in numerous species and in vitro, and none was found effective (Tzipori 1983). More recently, some possible treatment options have emerged for use in goats and other ruminants, notably halofuginone lactate and paromomycin sulfate. Neither drug is approved for use in goats in the United States.
Preliminary results from a French field trial indicated that the anticoccidial drug halofuginone lactate was effective in arresting diarrhea and eliminating shedding of oocysts in goat kids with cryptosporidiosis (Naciri et al. 1989). The drug was given orally at a dose of 0.5 mg/ kg daily for three to five days, and the therapeutic response was marked compared with affected kids in the same flock treated with sulfonamides. Since that time, the therapeutic efficacy of halofuginone in cryptosporidiosis has been more widely recognized, and the drug is now commercially available and approved for use in newborn calves in Europe for control of cryptosporidiosis. Available as an oral solution, it is given at a dose of 0.1 mg/kg bw for seven days, beginning on the first or second day of life. Field trials in goat herds in Greece showed that halofuginone lactate given orally to kids at a dosage rate of 0.1 mg/kg for seven days was effective in both the treatment and prevention of cryptosporidiosis, reducing the incidence of diarrhea, the duration and intensity of oocyst shedding, and death rates (Giadinis et al. 2008).
A cautionary note on the use of halofuginone is warranted. This drug has a very narrow margin of safety. It can be fatal when administered at three times the dose recommended. Animals should be weighed when possible to calculate an accurate dose and animals should be adequately hydrated before administration.
Paromomycin sulfate is an aminoglycoside antibiotic similar in structure to neomycin. It has been shown to have efficacy against C. parvum and is being used in human AIDS patients with cryptosporidiosis.
Several efficacy trials have been specifically carried out in experimentally infected (Mancassola et al. 1995) and naturally infected goats (Chartier et al. 1996a; Johnson et al. 2000). In all studies, the drug was administered orally to kids at a dose of 100 mg/kg bw but for varying durations of 10, 11, or 21 days. In all cases, there were marked reductions in oocyst shedding and clinical manifestations as compared to controls. When used for 21 days, in the face of a severe natural outbreak, treated animals showed no diarrhea and were negative for fecal oocysts on the day following cessation of treatment, while untreated controls in the same environment all developed diarrhea and had oocyst positive fecal samples (Johnson et al. 2000). Because the drug is given orally and is poorly absorbed from the alimentary tract, concerns about nephrotoxicity and ototoxicity associated with aminoglycoside administration, especially in diarrheic or dehydrated animals, are minimized.
There are also reports that decoquinate can reduce the severity of C. parvum infection and the number of oocysts shed in feces, though it does not prevent the development of clinical disease (Mancassola et al. 1997; Naciri et al. 1998; Ferre et al. 2005). In all these studies, the drug was administered orally to young kids for a period of 21 days at a dose of 2.5 mg/kg bw. In one study, however, the drug was also given to a group of pregnant does during their last three weeks of pregnancy, rather than to their kids. The positive effect on cryptosporidiosis in their kids was equivalent. The explanation was that treatment of the does suppressed the periparturient rise in oocyst shedding and therefore reduced environmental contamination and exposure of the newborn kids. Decoquinate is approved for use in the United States as an additive to feed or milk replacer for controlling coccidiosis in non-lactating goats, as discussed in the preceding section of this chapter on coccidiosis, but a higher dose is required for controlling cryptosporidiosis and such extralabel use of the drug is not permitted.
The non-reducing oligosaccharide alpha-cyclodextrin has also been evaluated as a treatment for cryptosporidiosis in goats. In an experimental model, kids given 500 mg/kg bw orally for six days beginning on the day of experimental infection had a longer prepatent period, a reduction in the patent period, a decrease in the intensity of infection, and far fewer cases of diarrhea compared with untreated neonatal kids (Castro-Hermida et al. 2004).
Control
The thrust of cryptosporidiosis control is in improved management and hygiene. Kids should be separated from does at birth and fed colostrum in clean nipple bottles. Goats that do not receive colostrum are more likely to develop cryptosporidiosis. Kids should be housed away from the adult herd, individually or in small groups. When kids are housed in groups, affected kids should be isolated at the first sign of diarrhea to reduce oocyst contamination of the environment. The author (DMS) has had excellent results in checking the spread of cryptosporidiosis in groups of young kids at risk by practicing early removal and strict isolation of affected kids at the first sign of diarrhea. Outbreaks can be stopped without the use of drugs.
Excellent sanitation in housing, bedding, and feeding utensils is probably most critical in controlling the spread of cryptosporidiosis among susceptible kids (Thamsborg et al. 1990). Rodent control may also be indicated, because there is evidence that oocysts derived from goats can infect mice, which in turn shed oocysts as well (Noordeen et al. 2002). In problem herds, water supplies should also be checked for contamination with C. parvum oocysts, because of the risk of water-borne infection (Watanabe et al. 2005). Personnel handling diarrheic kids should not handle healthy kids. Hands should be washed thoroughly after handling sick kids because of the zoonotic potential of cryptosporidiosis. Given that common disinfectants do not destroy cryptosporidia, steam cleaning or high-pressure cold water spraying of pens or hutches between uses is recommended. No vaccine is currently available, though this is an area of active research (Sagodira et al. 1999; He et al. 2002; Liu et al. 2010). While no drugs are specifically approved for prophylactic use in goats, published reports indicate that halofuginone, paromomycin, or decoquinate can help control the disease in young kids, as discussed above in the treatment section.