Diseases Caused by Bovine Viral Diarrhea Virus (BVDV)
Paul Walz
Definition and Etiology
Infections with bovine viral diarrhea virus (BVDV) are endemic worldwide, and BVD is recognized as one of the most important infectious diseases of cattle.
Not simply confined to causing disease in cattle and the alimentary system as the name would imply, BVDV is a pathogen capable of infecting other ruminants and pigs and affecting multiple physiologic systems, including the reproductive, respiratory, circulatory, immunologic, integumentary, musculoskeletal, and central nervous systems.1 Reproductive losses and respiratory tract disease are the most consequential economically, and infection of the reproductive system is critical for maintenance and transmission of BVDV within and among cattle populations through the generation of persistently infected (PI) offspring.2 Infection of pregnant cattle before day 125 of gestation can result in the birth of calves that are immunotolerant to and PI with BVDV.3 PI calves are the most significant reservoir of BVDV and are at risk for the sporadic, usually fatal mucosal disease.BVDV is the prototypic member of the genus Pestivirus within the Family Flaviviridae. The pestiviruses are enveloped, single-stranded, positive-sense RNA viruses. Historically, the genus Pestivirus contained four recognized species that include the two genotypes of BVDV (BVDV-1 and BVDV-2), classical swine fever virus and border disease virus. Genetically distinct pestiviruses that had not been designated as unique species included giraffe virus, Bungowannah virus, pronghorn virus, and HoBi-like viruses. The genus Pestivirus has undergone a change in nomenclature by the International Committee on Taxonomy of Viruses, with new species designations of Pestivirus A through Pestivirus K. The designations Pestivirus A through Pestivirus D correspond to the classic four species, BVDV-1, BVDV-2, classical swine fever virus, and border disease virus, respectively; the designations Pestivirus E through Pestivirus K correspond to pronghorn antelope pestivirus (E), Bungowan- nah virus (F), giraffe pestivirus (G), HoBi-like pestivirus (H), Aydin-like pestivirus (I), rat pestivirus (J), and atypical porcine pestivirus (K).4 A part of the logic for this new classification scheme is the knowledge that not all BVDV strains are host restricted to cattle.
As in many RNA viruses, mutations can occur within BVDV genomes and lead to genetic, antigenic, and pathogenic variation. BVDV exists as quasi-species, which are different but closely related mutant viral genomes subjected to continuous competition and selection. Nucleotide sequence differences are the most reliable criteria for differentiation of BVDV species. Subgenotypes of BVDV are described within BVDV-1 and BVDV-2 species: 21 among BVDV-1 viruses (1a to 1u) and 4 among BVDV-2 viruses (2a to 2d).5 Within the North American cattle population, there are three major subtypes, BVDV-1a, BVDV-1b, and BVDV-2a; the BVDV-1b subtype predominates in diagnostic laboratory submissions and prevalence studies of persistent infection.6 Predominance of the BVDV-1b strains in North America is noteworthy because most vaccines licensed and marketed in the United States contain BVDV-1a and BVDV-2 strains.7 Strains of BVDV can be also be further subdivided into cytopathic (CP) or noncytopathic (NCP) biotypes on the basis of their effect on cultured cells: CP strains cause vacuolation and death of cultured cells. The effect of the virus in cultured cells is not correlated with virulence, inasmuch as NCP BVDV are associated with cases of severe clinical disease, and only NCP strains of BVDV have been demonstrated, both naturally and experimentally, to induce persistent infection.8 The NCP biotype predominates in the cattle population, accounting for approximately 65% to 90% of BVDV isolates.9 The NCP biotype is often the source of CP strains, which arise by mutations and recombination in the NCP strain.
Epidemiology
PREVALENCE. BVDV is distributed in cattle populations throughout the world as indicated be serologic surveys that test for the presence of neutralizing antibodies to the virus. Exposure to PI animals or vaccination against BVDV can cause seropositivity in an animal within a population. Prevalence of seropositive cattle varies among countries, with differences in seroprevalence rates largely dependent on management of cattle populations examined.
Use of vaccines, open versus closed cattle production units, population densities, indoor versus pasture housing, communal grazing, and presence of PI animals greatly influence the numbers of seropositive cattle. Surveys in North America have indicated high seropositive rates in cattle, approaching 90%.10 In contrast, the number of PI cattle is much lower and is generally believed to account for fewer than 1% of all cattle. In randomly selected beef herds in five U.S. states, the prevalence rates of BVDV persistent infection in individual animals and in herds were 0.17% and 2.7%, respectively.11 Other persistent infection prevalence studies in beef cow-calf operations within more geographically focused areas in the United States have demonstrated rates between 0.09% and 1.7%, respectively12,13; however, not all studies performed have involved randomly selected herds, which can affect prevalence rates considerably. Surveys conducted on United States dairy operations have demonstrated individual animal prevalence rates of 1.9%14 and 0.13%.15 PI cattle can be found in clusters within groups of cattle as a result of BVDV exposure in cows at similar times in pregnancy, and this can elevate the prevalence within a studied population. Although it is generally believed PI calves have poor survivability, they can survive to adulthood, become pregnant, and give birth to PI offspring. Of PI calves identified from randomly selected beef herds, 61% survived to weaning and thus were capable of providing a source of virus throughout the breeding season and early gestation.11 The numbers of PI calves arriving at United States feedlots represent a prevalence rate between 0.1 and 0.4%.16,17HOST RANGE. Cattle are considered the natural host of BVDV; however, BVDV is not strictly specific about hosts.18 Historically, pestiviruses were assigned according to the species from which they were isolated; BVDV, classical swine fever virus, and border disease virus isolates were recovered from cattle, pigs, and sheep, respectively.
Through serologic evidence, BVDV infections have been identified in more than 50 species within 7 of the 10 families of the mammalian order Artiodactyla. Species other than cattle that have been reported to be susceptible to BVDV infection include pigs, sheep, goats, bison, captive and wild cervids, and Old World and New World camelids.18 Of the heterologous hosts, sheep and goats are considered to have high potential for BVDV infections because they are phylogenetically most closely related to cattle, and sheep and goats can have close contact with cattle under certain management conditions such as subsistence agriculture and hobby farming. Alpacas and white-tailed deer have received attention as heterologous hosts for BVDV infections because of the identification of PI animals. PI alpacas can survive for several months, but low birth weights, failure to thrive, and chronic respiratory and gastrointestinal infections are described in such animals.19 The BVDV-1b strain is most frequently isolated from PI alpacas and could be the result of introduction and intraspecific spread and maintenance of BVDV1b into North American alpaca populations; an alternative explanation is that unique BVDV1b subgenotypes are able to establish transplacental infections in alpacas.20Infection of wildlife with BVDV has been demonstrated by several serosurveys and, less frequently, virus identification in individual animals. The livestock-wildlife interface is of great concern for a number of infectious diseases, including the related classical swine fever virus in wild boars. Wildlife may become infected with BVDV, but other factors, including shedding of the virus, intrapopulation maintenance, and amount of interspecific contact may influence the establishment of BVDV wildlife reservoirs. Persistent infections of white-tailed deer with BVDV have been demonstrated both by experimental and natural exposure.21 As in cattle, the most dramatic effects of BVDV infections in white-tailed deer involve the reproductive system.
Shedding of BVDV through nasal or rectal routes has been demonstrated in acutely infected and PI white-tailed deer, and shedding has resulted in transmission to other white-tailed 2223deer.22, Apparent prevalence rates of persistent infections in United States cervid populations were between 0.03 and 0.3% in tested populations.24-26 Whether BVDV infection in these populations originated from contact with cattle or was the result of an endemic cycle is unknown.
Identification of heterologous PI hosts may have important implications for the epidemiologic behavior of BVDV; of most importance is that nonbovid PI animals may serve as reservoirs for BVDV As reservoirs, nonbovid PI animals could threaten the health and well-being of other artiodactyls, including cattle, and adversely affect outcomes and ultimate success of organized BVDV control and eradication efforts.27
TRANSMISSION. Methods of BVDV transmission can be classified as vertical and horizontal. With BVDV, horizontal and vertical transmissions are closely related, as horizontal transmission to a pregnant cow can result in vertical transmission to the developing fetus (Fig. 32.91). As stated previously, PI cattle can reach adulthood. Calves delivered from PI dams are themselves PI; however, the status of most PI offspring is the result of an acute infection, or horizontal transmission, in the dam.11 BVDV can be horizontally transmitted continuously by PI animals and briefly by animals during an acute infection. As with other viral diseases, factors controlling BVDV transmission in a susceptible population are the infectiousness of the virus, the contact rate between infectious and susceptible animals, the duration of infectiousness, and the proportion of the population that is naive and susceptible.28 Cattle PI with BVDV shed large amounts of virus their entire life and are the major source of BVDV transmission both within and among herds.
They are capable of shedding virus from nasal secretions, urine, feces, milk and colostrum, uterine fluids, saliva, tears, and semen.29 Direct contact with these body fluids from PI cattle is the most efficient method of horizontal BVDV transmission, inhalation and ingestion being the most common routes. Although heterologous PI hosts are capable of BVDV in nasal secretions in amounts equivalent to those shed by PI cattle,21 they are assumed to play a reduced role in BVDV transmission because rates of contact rates between infectious heterologous hosts and susceptible cattle are lower.Acutely infected cattle also have a limited role in BVDV transmission because levels and duration of virus shed are considerably lower; however, potential for transmission of BVDV after acute infection can be greatly influenced by the virulence of the acutely infecting strain of BVDV, the proportion of naive animals being exposed, and the immune cross-protection acquired through either vaccination or exposure to field strains of virus. Calves acutely infected with a highly virulent strain of BVDV shed greater amounts of virus and for a longer duration,30 and so they theoretically are a source of transmission. However, an experimental study demonstrated no spread of infection from acutely infected to susceptible animals even though they were in close contact, which further emphasizes the importance of virus and host characteristics during
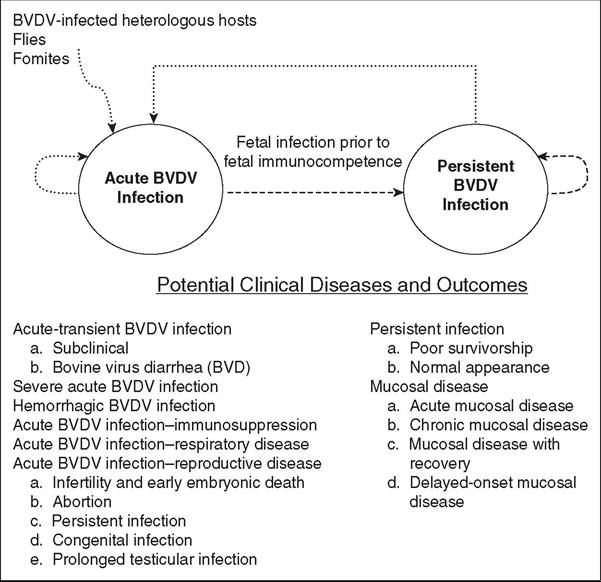
FIG. 32.91 Two forms of bovine virus diarrhea virus (BVDV) infection: acute and persistent. Routes of horizontal transmission are indicated by dotted lines (most consequential is from persistently infected [PI] cattle to naive cattle); routes of vertical transmission are indicated by dashed lines (most consequential is acute BVDV infection in pregnant cattle). Potential clinical diseases are not exclusive, and multiple clinical forms can be observed in the same population of cattle.
transmission.31 After acute BVDV infection, virus can be found in nasal secretions usually for only a few days between 4 and 10 days after exposure.
Although contact with PI cattle is the most widespread cause of BVDV transmission, infections with BVDV in the absence of PI cattle have been described in seronegative herds.32 Spread of BVDV can occur in the absence of PI cattle as a result of direct contact with acutely infected animals, contact with heterologous hosts, or indirect contact. Because BVDV is an enveloped virus, it is unstable at low or high pH and at high temperatures but can survive in a cool, protected environment for several weeks.29 Temperature has a major influence on BVDV survival in manure slurry: The virus can remain infective for more than 6 weeks at 5° C but less than 2 weeks at 20° C.33 According to research with the related classical swine fever virus, BVDV is probably susceptible to common disinfectants such as chlorhexidine, phenols, iodophors, aldehydes, and hypochlorites.34 Indirect transmission through contaminated needles and equipment and through insect vectors has been demonstrated experimentally.35,36 Insects such as stable flies, horse flies, and face flies can serve as potential sources of BVDV infection35,37; however, the horn fly, a common fly species affecting cattle, cannnot transmit BVDV between PI cattle and BVDV-naive cattle.38
Bulls acutely infected and PI with BVDV are capable of shedding virus in semen and have the potential for horizontal and vertical transmission.39-41 The virus may survive cryopreservation and processing of semen for artificial insemination.42 Although acutely infected bulls shed lower concentrations of BVDV in semen than do PI bulls, infection of artificially inseminated heifers may result from the use of semen from acutely infected bulls before seroconversion.40 Prolonged, localized testicular infections have also been described after experimental BVDV infection of postpubertal bulls; however, their prevalence and epidemiologic significance are probably low.43
Herd-to herd transmission usually occurs through acquisition of PI cattle or pregnant cattle that are carrying PI fetuses. Introduction of pregnant adult females carrying PI fetuses, aptly named “Trojan dams,” have been recognized as an important source of infection transmission to BVDV-free herds.44 Cattle operations that have purchased cattle within the past 5 years are at highest risk for having PI animals.45 Purchase of new cattle incubating an acute infection or exposure to other cattle through fence line contact, communal pastures, and animal exhibitions can also enable BVDV introduction into a herd.
Clinical Signs and Differential Diagnoses
BVDV infection in cattle may result in a complex spectrum of clinical manifestations ranging from subclinical to fatal disease. The virus is capable of infecting multiple organ systems, often concurrently. Clinical manifestations are dependent on the interplay of host factors, environmental stress levels, and viral factors.46 Host factors influencing the outcome of viral infection include immune status, pregnancy status, gestational age of the fetus at the time of infection, and whether the host is immunotolerant or immunocompetent. Viral factors include biotypic variation, genotypic variation, and antigenic diversity. With regard to the clinical manifestations of BVDV infection in cattle, both BVDV-1 and BVDV-2 strains can be involved in the entire spectrum of clinical disease, including acute and persistent infections. Two states of infection (acute and persistent) are possible with BVDV infections in ruminants, and this is the basis for discussion of clinical signs and pathogenesis within this text (see Fig. 32.91).
Disease Associated With Acute BVDV Infection
The terms acute, primary, and transient have all been used to describe BVDV infection in postnatal cattle. Although postnatal cattle are able to respond immunologically to BVDV, immunosuppression induced the by virus is an important contribution to pathogenesis and clinical disease. For discussion of clinical disease states, acute BVDV infections can be further subdivided into six disease categories: acute-transient BVDV infection, severe acute BVDV infection, hemorrhagic BVDV infection, acute BVDV infection/immunosuppression, acute BVDV infection/respiratory disease, and acute BVDV infection/reproductive disease.
ACUTE-TRANSIENT BVDV INFECTION. Acute-transient BVDV infection occurs in postnatal cattle. In most situations, the disease occurs in cattle considered BVDV naive, but disease could potentially occur in cattle seropositive for BVDV that become infected with a heterologous BVDV strain. Most acute BVDV infections occur without clinical manifestations; up to 90% of BVDV infections are estimated to occur without clinical signs.46 If cattle are observed closely, however, mild fever, leukopenia, and a decrease in milk production can be observed. Exposed cattle develop BVDV-specific neutralizing antibodies. If a pregnant animal has even subclinical BVDV infection, deleterious effects can be manifested in the fetus.
Bovine viral diarrhea is the term used to describe the clinical form of BVDV infection in immunocompetent cattle, and digestive tract disease manifested by severe diarrhea was part of the original description of BVD in 1946.47 The incubation period for acute BVDV in naive cattle is 5 to 7 days. After oronasal uptake, BVDV initially replicates in nasal mucosa and tonsils. Viremia typically lasts for several days starting around day 3 after infection, but it can last for up to 15 days. During this period of viremia, spleen, thymus, lymph nodes, gastrointestinal epithelium, and the respiratory tract can become sites of further viral replication. Clinical signs of BVD include depression, oculonasal discharge, anorexia, fever, oral erosions and ulcerations, diarrhea, and, in lactating cows, decreased milk production. The respiratory rate may be rapid, which may be interpreted incorrectly as pneumonia. Clinical pathologic changes such as leukopenia are observed in calves acutely infected with BVDV.46 The major decreases in the total leukocyte count have been in the numbers of neutrophils and lymphocytes.
One of the most important determinants for the outcome of acute BVDV infection in susceptible animals is the virulence of the infecting BVDV strain.48 Experimental infections with BVDV strains of low and high virulence reveal similar initial infection and patterns of dissemination through the body but differences in the amount of virus in tissues and in speed of dissemination.49 Virulent strains of BVDV result in widespread distribution of virus in tissues, and BVDV strains that induce high levels of viremia result in more severe clinical signs. Viral replication and antigen distribution can be detected in the thymus, lymph nodes, Peyer's patches, tonsils, and spleen with all strains of BVDV. In contrast to strains of BVDV with low virulence, in which antigen is found only outside of lymphoid tissues in the intestinal mucosa, antigen against high virulent strains can be found in bone marrow, intestinal mucosa, respiratory tract, endocrine tissues, and nervous system tissues after acute infection.49 The HoBi-like viruses have received considerable attention because of their emergence and detection in South America, Europe, and Southeast Asia, and natural descriptions and experimental reproductions of infection with HoBi-like viruses have revealed very mild disease signs in cattle. Most description of HoBi-like virus infection in natural settings are related to either respiratory disease or reproductive disease, specifically abortion. During experimental reproduction of the disease, clinical signs associated with HoBi-like virus infection can be indistinguishable from those of BVDV infection with 50
strains of low virulence.50
Disease associated with acute BVDV infection is often observed in seronegative cattle between the ages of 6 to 24 months. It is generally believed that maternally derived BVDV- specific antibodies have waned and acquired immunity is not yet formed, and thus calves as young as 4 to 6 months of age are susceptible. Maternally derived BVDV-specific colostral antibodies are thought to be protective unless sufficient antigenic diversity exists between the challenge strain and strain against which the maternally derived colostral antibodies were developed. Disease as a result of acute infection can be observed in preweaned calves.51 Acute disease in neonatal or preweaned calves can result in signs of pneumonia or enteritis, often in association with failure of passive transfer. In studies of the duration of maternal colostrum-derived antibodies against BVDV-1 and BVDV-2, a major finding is the variability in duration between calves as a result of colostrum quality and consumption. Estimated mean time to reach seronegative status in dairy calves receiving maternal colostrum was 117.7 ± 37.7 days for BVDV-1 and 93.9 ± 61.9 days for BVDV-2.52 In another study, the mean duration for colostrum-derived antibodies in beef calves was 185.6 ± 59.8 days for BVDV-1 and 157.8 ± 56.1 days for BVDV-2.53
Diarrheal diseases considered in the differential diagnosis for acute BVDV infection in adults include salmonellosis, winter dysentery, paratuberculosis, intestinal parasites, MCF, arsenic poisoning, and copper deficiency (molybdenosis). The differential diagnoses for acute BVDV infection for the neonatal period includes other causes of diarrhea in young calves, such as rotavirus or coronavirus infection, cryptosporidiosis, enterotoxigenic E. coli infection, salmonellosis, and coccidiosis. Other causes of calf pneumonia such as bovine respiratory syncytial virus, pasteurellosis, hemophilosis, or mycoplasma infection should also be considered.
SEVERE ACUTE BVDV INFECTION. An atypical and significantly more severe form of BVDV infection was first described in cattle in the late 1980s.54 The disease had a peracute course, caused morbidity in high rates, and resulted in substantial numbers of deaths in all age groups. In Quebec, mortality due to BVDV in veal operations was estimated at 25% of 143,000 calves for 1993.55 The outbreak in Ontario involved 150 dairy, 600 beef, and 100 veal herds. The severity of disease varied among herds; in some herds, mortality rates were 10 to 20% in all age groups of cattle. Clinical manifestations of severe, acute BVDV infections include diarrhea, pyrexia, decreased milk production, and, frequently in older cattle, oral ulcerations. Because of the presence of severe clinical disease in combination with oral ulcerations, mucosal disease was a primary part of the differential diagnosis in the late 1980s and early 1990s. Other diseases that cause oral lesions in cattle include MCF, vesicular stomatitis, papular stomatitis, FMD, and bluetongue. Abortions in cattle also were common. These outbreaks of severe clinical disease were reported to be associated with the introduction of cattle to the herd or history of inadequate vaccination programs.54 BVDV isolates from outbreaks in Canada and the United States were analyzed, and NCP isolates were obtained from affected cattle. Nucleotide sequencing of the RNA genome of these isolates revealed a distinct group of BVDV, which were designated as BVDV-2,56 which is now considered a separate species within the genus Pestivirus.
The outbreaks of severe BVDV infections clearly demonstrated that some BVDV strains can cause severe life-threatening disease in postnatal, immunocompetent cattle. Before 1993, it was believed that the majority of BVDV infections in immunocompetent, adult cattle resulted in subclinical or mild disease. Although BVDV-2 strains were associated with severe acute BVDV infections, not all BVDV-2 strains cause severe disease. Furthermore, it is likely that some BVDV-1 strains are capable of inducing severe clinical disease under unfortunate conditions involving host, agent, and environment factors.
HEMORRHAGIC BVDV INFECTION. The hemorrhagic syndrome, which is characterized by thrombocytopenia, is an additional form of severe acute BVDV infection in immunocompetent cattle.57 Hemorrhagic syndrome was first described in both calves and adult cattle naturally infected with BVDV; severe depressions in platelet counts and high case fatality rates were reported.58 Clinical manifestations of the hemorrhagic syndrome are related primarily to thrombocytopenia and include bloody diarrhea, epistaxis, petechial hemorrhages, ecchymotic hemorrhages, hyphema, and bleeding from injection sites or insect bites. Fever, diarrhea, rumen stasis, and dehydration are also common. In addition to thrombocytopenia, leukopenia characterized by severe neutropenia and lymphopenia are clinical pathologic abnormalities. Thrombocytopenic BVDV infections have been experimentally reproduced, almost exclusively with NCP BVDV-2 strains57,59; however, hemorrhagic syndrome has been reported after experimental inoculation of colostrum-deprived calves with a BVDV-1b strain.60 Platelet dysfunction has also been described with experimental BVDV infection61; thus quantitative and qualitative platelet defects contribute to the hemorrhagic diathesis observed in infected cattle. Bone marrow infection, specifically BVDV infection in bone marrow megakaryocytes, may be important in the origin of BVDV-induced thrombocytopenia.30,62 Diseases that can mimic hemorrhagic syndrome include septicemia with subsequent development of disseminated intravascular coagulation, sweet clover poisoning, and bracken fern poisoning.
acute BVDV Infectionzimmunosuppression. Although the majority of BVDV infections in immunocompetent cattle are transient and self-limiting, it is apparent that when infection occurs in the presence of other microorganisms, BVDV can contribute to disease that becomes clinically evident. Numerous studies, under both natural and experimental conditions, have demonstrated the relationships between BVDV and other infectious agents. The virus has been reported to enhance the pathogenicity of bovine herpesvirus-1 (BoHV-1), actinomycosis, papular stomatitis, enteritis caused by Salmonella spp., colibacil- losis, metritis, and mastitis. The role of BVDV in synergistic or mixed infections with other agents is probably to cause immunosuppression.63 Mechanisms responsible for BVDV- induced immunosuppression are probably multifactorial; however, critical information on mechanisms of the suppression, the duration and extent of recovery, and the long-term effects are undetermined.27 The ability of BVDV to induce quantitative and qualitative defects in leukocytes could provide some contribution to immunosuppression.
The defects involve immune system cells of both the innate and adaptive immune responses. A transient leukopenia occurs in most cattle acutely infected with BVDV that may be more severe with BVDV-2 than with BVDV-1. Decreases in helper (CD4+), cytotoxic (CD8+), and gamma/delta T-lymphocytes, as well as B-lymphocytes and neutrophils, have been reported and these decreases are often strain dependent.64 The mechanism of leukopenia has been explained in several possible ways; these include immune system removal of BVDV-infected immune cells, destruction of immune cells by BVDV, and increased trafficking of immune cells into tissue sites of viral replication. During acute infection, various levels of lymphoid depletion are observed in the thymus, spleen, lymph nodes, and gut-associated lymphoid tissues (Peyer's patches).65 The NCP BVDV strains are lymphotrophic, and lymphocyte apoptosis in lymphoid tissues results in cytolysis and lymphoid depletion.
Decreased functional responses in immune system cells have also been described during acute BVDV infection, and affected cells include lymphocytes, neutrophils, monocytes, and mac- rophages.66,67 Dysfunction in both B and T cells has been demonstrated by poor culture response to phytohemagglutinin, pokeweed mitogen, and concanavalin A stimulation and by decreased production of interferon.68 Infection of peripheral monocytes in vitro caused a significant decrease in the chemotactic responses to lymphokine.69 Immunohistochemical and flow-cytometric studies have demonstrated that BVDV infects and replicates in antigen-presenting cells.70 Infection of monocytes with BVDV has been shown to stimulate the synthesis of cytokines that may reduce their ability to stimulate T cell responses to specific antigens and mitogens. A decrease in the secretion of TNF-α from LPS- or Salmonella-stimulated bone marrow-derived macrophages that had been infected with BVDV has been reported.71 These abnormalities, in combination with environmental or management stressors, or both, contributes to immunosuppression and concurrent infection with other pathogens. Increased susceptibility to other secondary infections is a consequence of BVDV-induced immunosuppression.
acute BVDV Infectionzrespiratory disease. Substantial data demonstrate that BVDV infection plays an important role in polymicrobial disease, and the bovine respiratory disease complex in feedlot animals and intensively housed calves is the best example of this type of disease process. In North American feedyards, bovine respiratory disease is the most frequently reported cause of morbidity and mortality. The role of BVDV in the bovine respiratory disease complex is debatable as to whether BVDV-induced immunosuppression or primary infection of the respiratory tract plays the major role, but BVDV can be isolated from pneumonic cattle more frequently than other viruses.72 Certain BVDV strains may be pneumotrophic, as evidenced by some BVDV strains that cause interstitial pneumonia in experimentally inoculated calves.73
BVDV may play a secondary role in bovine respiratory disease complex through immunosuppression. Although BVDV is frequently isolated from cattle with pneumonia, it is often isolated with the other infectious agents typically associated with bovine respiratory disease complex. Reports from disease outbreaks and experimental studies have supported a synergistic role of BVDV with BoHV-1 and bovine respiratory syncytial virus.74,75 Calves experimentally inoculated with BVDV 7 days before inoculation with BoHV-1 developed more severe clinical disease, with dissemination of BoHV-1 into nonrespiratory tissues such as intestinal and ocular tissues, than did calves inoculated with BoHV-1 alone.74 Initial BVDV infection may impair the ability of calves to clear BoHV-1 from the lungs and to contain BoHV-1 at the local infection site. Synergism between BVDV and bacterial pathogens, specifically Mannheimia haemolytica and Mycoplasma bovis, has also been described.76,77 Viral-bacterial synergy in bovine respiratory disease complex probably is the result of BVDV compromise of the host defense mechanisms. Viral damage of nasal turbinate and tracheal epithelium alters mucociliary clearance of bacteria with subsequent bacterial attachment and colonization. The innate defenses of the lung are compromised with BVDV infections of macrophages and neutrophils, which results in depletion, destruction, or impairment in function. B and T cell-mediated immune responses can also be suppressed as a result of BVDV infection.63
acute BVDV Infectionzreproductive disease. Reproductive tract infections have been described for male and female cattle. The importance of BVDV on the male reproductive tract has not received attention equivalent to the effects on female reproduction for a number of reasons, but from a transmission standpoint, acutely infected bulls and PI bulls are capable of shedding virus in semen.39 Either acceptable or unacceptable semen quality may be observed in bulls acutely infected with BVDV. Although acute BVDV infections generally result in a transient viremia with subsequent clearance of the virus by the host immune system, a localized, prolonged infection of testicular tissue with NCP BVDV has been described in both natural and experimental conditions.39,41 The original description of prolonged testicular infection with BVDV was identified in the testes of a seropositive, nonviremic bull at an artificial insemination center.41 Despite absence of viremia, the bull continuously shed infectious BVDV in semen throughout his life, and the semen of this bull resulted in infection and subsequent seroconversion of an inseminated seronegative heifer.78 Furthermore, this bull displayed a consistently high concentration of circulating serum antibodies that neutralized the specific viral strain that was persistently shed in the semen. Since that original description, prolonged testicular infections with BVDV have been reproduced in post-pubertal bulls. Viral RNA has been detected in semen for 2.75 years after BVDV exposure, and infectious virus grown from testicular tissue has been detected up to 12.5 months after BVDV exposure.39 Protection from a systemic immune response due to a bloodtestes barrier is believed to be the mechanism for the localized, prolonged testicular infection.
Infection with BVDV can cause significant reproductive problems in nonimmune pregnant cattle, including fertilization failure, embryonic death, abortion, persistent infection, and congenital defects (Fig. 32.92). Transplacental infection is a common event in pregnant cattle exposed to BVDV, and in experimental studies, transplacental infection occurs with almost 100% efficiency.79 As discussed previously, most acute BVDV infections are subclinical, but infection in pregnant cattle may result in significant disease. The economic damage caused by BVDV in susceptible herds is associated mainly with the outcomes of intrauterine infections. The outcome of fetal infection with BVDV is dependent on three main factors: (1) time of infection (i.e., gestational age of the fetus at the time of infection); (2) organ system involved in the infection; and (3) properties of the virus (i.e., biotype, virulence, and target cell range). Of importance is that BVDV-1 and BVDV-2, as well as CP and NCP strains, can result in reproductive disease, although only NCP strains are capable of resulting in PI cattle.
Infertility and Early Embryonic Death. Infection of cattle before insemination can result in impaired conception rates.80 This may be due in part to ovarian infection and dysfunction as a result of BVDV viremia. Viral antigen in macrophagelike cells and ovarian stromal cells and oophoritis have been
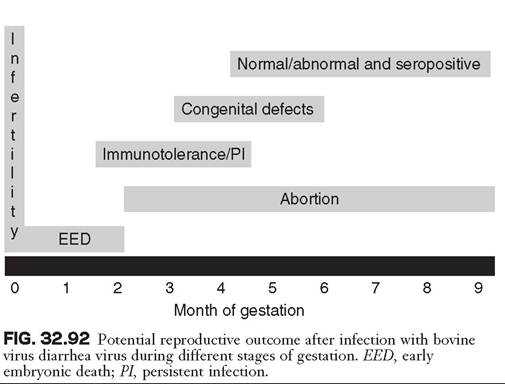
described in cattle acutely infected with NCP BVDV81 and in cattle vaccine with modified live-virus vaccines containing CP BVDV.82 Of particular interest is that pathologic change and BVDV antigen were detected in some cattle up to 60 days after infection.83 The first 45 days of gestation are the embryonic period, and conception and pregnancy rates are lower if cattle are exposed at the time of insemination and become viremic during the first 3 weeks of pregnancy.
Abortion. BVDV infection can cause abortion at any time during gestation. Fetal resorption, mummification, or expulsion can occur after fetal death, depending on the gestational age of the fetus at the time of infection.84 Although fetal death and abortion are most common during the first trimester, material from middle- and late-term abortions and stillbirths should be examined for BVDV. Diagnostic laboratory surveys indicate that BVDV isolation prevalence from aborted animal fetuses in the United States is quite variable; BVDV has been isolated in as few as 0.1% and as many as 27.2% of submitted fetuses.85,86 After BVDV infection, expulsion of the fetus may occur shortly or may be delayed for several months after infection. As a result of delayed expulsion after fetal death, isolation of BVDV from aborted fetuses may not be successful. This has also been demonstrated experimentally, whereby abortion was observed 30 to 50 days after experimental infection at approximately days 100 to 120 of gestation.87
Persistent Infection in Offspring. Gestational days 45 to 125 are the period of fetal development during which fetal immunocompetence for BVDV develops. The most outstanding outcome of infection during this period is the development of persistent infection. Immunocompetence for BVDV is not absolute at day 125 of gestation, and the dams of most PI calves were exposed before gestational day 100. By definition, cattle that are PI with BVDV are immunotolerant to BVDV. Biotype plays an important role in infection during this period of gestation. Abortion can occur with infection with both CP and NCP strains of BVDV; however, only NCP strains are capable of causing persistent infection.8 The PI animal's body cannot recognize the infecting NCP BVDV as foreign. Immunotolerance is specific to the infecting NCP strain of BVDV, and after birth, PI animals can respond immunologically to heterologous strains of BVDV. For this reason, PI animals may be seropositive for BVDV, and seropositive status cannot be utilized diagnostically to rule out persistent infection.
Congenital Defects. Congenital malformations are associated with BVDV infection during the gestational period of organogenesis, which occurs approximately between days 100 and 150 of gestation.79 Congenital malformations that have been reported in association with BVDV infection include cerebellar hypoplasia, hypomyelinogenesis, hydranencephaly, alopecia, cataracts, optic neuritis, brachygnathism, hydrocephalus, microencephaly, thymic aplasia, hypotrichosis, pulmonary hypoplasia, and growth retardation. Congenital malformations may be due to the direct result of viral infection on developing cells, or the host immune response in destroying virally infected cells. During a point-source of exposure of BVDV into a bred population of cattle, congenitally malformed calves will be born just before the birth of PI animals, and this information can be useful for determining sampling of calves for BVDV.
BVDV Infection During Later Stages of Pregnancy. Fetal infections can occur in the last trimester, but at this stage of gestation, organogenesis is often complete and fetuses are considered immunocompetent to BVDV. Congenital infection during this period usually results in the birth of a calf that is seropositive on a precolostral blood sample. Detrimental effects of BVDV infection in late gestation seem to be less common; however, weak calves and abortions have been reported.79 It has generally been accepted that infection in late gestation results in the birth of calves that are clinically normal; however, calves congenitally infected with BVDV may be at greater risk for increased morbidity and mortality postnatally.88 The congenitally infected calves that are born with neutralizing antibody titers to BVDV have been shown to be twice as likely to experience a severe illness during the first 10 months of life as are calves born free of BVDV antibody titers.88 The prevalence and importance of congenital infections remains to be determined.
Disease Associated With Persistent
BVDV Infection
PI cattle are the main reservoir for BVDV Cattle PI with BVDV are characterized by a wide distribution of BVDV throughout their organs as a result of lacking an immune response to the persisting BVDV strain. In PI animals, the virus is present throughout all organs and tissues; therefore BVDV is shed in multiple body excreta, including nasal discharges, urine, semen, colostrum/milk, and feces. Some PI calves are born weak and stunted and die shortly after birth46; however, many PI calves are born without any observable abnormalities and can survive for at least several months. This is important for beef cattle herds because these animals can survive into the start of the subsequent breeding period and thus become an important source of horizontal transmissions. PI cattle can have decreased survival, as evidenced by reports documenting a mortality rate higher than 50% in the first year of life.89 This mortality may be due in part to the fact that PI cattle may have an impaired immune response, which makes them more susceptible to opportunistic pathogens. The rate of vertical transmission is 100% as PI cows give birth to PI offspring.
PI cattle are at risk of developing mucosal disease. Mucosal disease is the most dramatic of BVDV-associated clinical disease because of the severity and characteristics of lesions. Mucosal disease occurs when cattle that are immunotolerant and PI with NCP BVDV become infected with a CP BVDV.90 Because mucosal disease occurs only in PI cattle and PI cattle are less than 1% of the cattle population, it is characterized as a disease with low case attack rate and high case fatality rate. Although the origin of the CP BVDV can be external, such as after the use of modified live-virus vaccines, current theory holds that in the majority of cases of mucosal disease, the CP BVDV represents a mutation of the NCP BVDV (the persistently infecting biotype).90 There are multiple clinical forms of mucosal disease; the variations are due to the antigenic relationship between the persistently infecting NCP strain and the superinfecting CP strain.90 The multiple clinical forms of mucosal disease can be categorized as acute (fatal) mucosal disease, chronic mucosal disease, chronic mucosal disease with recovery, and delayed-onset mucosal disease.90
ACUTE MUCOSAL DISEASE. Acute mucosal disease occurs when the superinfecting CP strain shares close antigenic homology to the PI NCP strain. The CP strain may arise de novo by a mutational event from the NCP strain in the PI animal. The mutational event affects the viral biotype but does not affect the viral antigenicity.91 The clinical findings associated with acute mucosal disease include pyrexia, depression, anorexia, decreased milk production, profuse diarrhea, ptyalism, mucopurulent oculonasal discharge, and oral erosions.46 Oral erosions are present on the lips, gingival margins, tongue, dental pad, and the hard palate. Experimental reproduction of acute mucosal disease indicates an incubation period of 7 to 14 days.90 Acute mucosal disease is considered 100% fatal.
CHRONIC MUCOSAL DISEASE. Chronic mucosal disease and mucosal disease with recovery are forms of mucosal disease in which the superinfecting CP strain is heterologous to the PI NCP strain. The likely source of the superinfecting CP strain is external, rather than a mutational event in the PI NCP strain. The clinical manifestations of chronic mucosal disease are anorexia, weight loss, diarrhea, chronic bloat, alopecia, erosive lesions on the mouth and skin, and lameness.46 The lameness may develop as a result of laminitis, interdigital necrosis secondary to erosive lesions, and hoof deformities. In rare cases, a PI animal may mount an immune response to the superinfecting, heterologous CP strain and clear the superinfection; this situation has been referred to as mucosal disease with recovery.90 Although recovery may be possible, a PI animal will always remain PI with the NCP strain.
Necropsy Findings
Pathologic lesions associated with BVDV infection can be quite diverse in types and degree and, like the clinical manifestations, are dependent on host and viral factors. Age of animal, immune status at the time of infection, and the presence of concurrent infections greatly influence the pathologic observations. Many lesions observed are not specific to BVDV infection; thus laboratory testing becomes an important component in the diagnostic process. Occasionally, only microscopic lesions are present, and unless histopathologic study is performed, a lesion can be unnoticed and infection undiagnosed.
ACUTE BVDV INFECTION. Animals that die of acute BVDV infection have less severe lesions. In addition, lesions, when present, can often be nonspecific and subtle. Edematous intestines, occasionally with ecchymotic hemorrhages and mild reaction in mesenteric lymph nodes, can be observed. Occasionally, linear erosions in the alimentary tract and proximal colon are present, along with atrophy of gut-associated lymphoid tissues (Peyer's patches). Secondary bacterial infections such as enteritis, pneumonia, or mastitis are present and have contributed to death. Animals that die of hemorrhagic syndrome can have evidence of hemorrhage in many organ systems, including the gastrointestinal, cardiovascular, respiratory, and urinary systems. Petechial or ecchymotic hemorrhages are often apparent on mucosal surfaces, and hemorrhage in the gastrointestinal submucosa and Peyer's patches may be significant. Lesions associated with cases of severe acute BVDV infection can be similar to lesions associated with mucosal disease. Because of this and the severe clinical signs observed, it is usually not possible to differentiate the two forms of disease without supporting laboratory testing.
MUCOSAL DISEASE. Postmortem examination of cattle dying of mucosal disease usually reveals gross abnormalities most prominent in the alimentary system. Animals that die of mucosal disease usually have severe, necrotizing erosive or ulcerative lesions involving the mouth, tongue, esophagus, ruminal pillars, omasum, abomasum, intestines, and cecum.92 Erosive lesions may extend onto the external nares and into the nasal cavity. Ulcers involving the esophagus typically are linearly elongated. Peyer's patches in the small intestine often are necrotic and hemorrhagic. Bowel contents are watery, hemorrhagic, and foul smelling. Pathologic gastrointestinal conditions may be absent or very mild in cattle with chronic mucosal disease, although histopathologic lesions usually are present with atrophy of Peyer's patches. Skin lesions are often found and include patchy hyperkeratosis around the neck, shoulder, and perineal areas. Erosive lesions involving the perineal area, the prepuce, and the interdigital cleft and coronary band of the hoof may be present. Skin lesions are most apparent in cattle suffering from chronic mucosal disease.
ABORTIONS AND CONGENITAL DEFECTS. Pathological criteria for the diagnosis of BVDV as a cause of abortion are not established. Fetuses aborted as a result of in utero BVDV infection often are autolytic when expelled. Lesions found in aborted fetuses and the accompanying placentas are nonspecific for BVDV Under experimental conditions or when aborted fetuses are expelled soon after death, lesions observed include conjunctivitis, peribronchiolar and interalveolar pneumonia, and nonspecific myocarditis. A significant decrease in cerebellar mass may be evident in calves affected by cerebella hypoplasia as a result of in utero infection with BVDV during the second trimester of gestation. Other common congenital defects associated with BVDV infection that may be noted at necropsy include cataracts and thymic hypoplasia.
Diagnosis
Numerous methods are available for detecting BVDV infection. The choice of test depends on the clinical problem, the availability of tests by different diagnostic laboratories, and financial considerations (Table 32.12). Because the clinical manifestations of BVDV are diverse, accurate diagnosis of BVDV infection relies on laboratory testing. Sample selection and handling for BVDV testing is critical because the initial test material largely defines success or failure of the diagnostic test.93 The introduction of PCR has transformed viral diagnostics, and real-time PCR technology has further improved the already significant benefits of traditional PCR. Real-time PCR has become the tool often used by diagnostic laboratories for the detection of BVDV.
VIRUS ISOLATION. Virus isolation is a basic, reliable, and widely used method for diagnosis of BVDV infection, and remains the “gold standard” of BVDV diagnostic tests. The BVDV may be cultured and isolated from many samples, including serum, whole blood, semen, nasal swabs, and various types of tissues.94 Although their use is time consuming and expensive, mononuclear cells in the buffy coat from whole blood are an ideal sample for individual virus isolation when an acute BVDV infection is suspected because BVDV is highly cell associated and neutralizing antibodies to BVDV in serum do not interfere with the virus isolation.94 Sample handling is very important for virus isolation with mononuclear cells because they need to be viable when inoculated in the cell culture.93 The ideal tissue samples for virus isolation from deceased animals at postmortem examination include lymphoid organ- related tissues such as spleen, Peyer's patches, mesenteric lymph nodes, and thymus.94 In addition, pooled tissues from the same animal can be applied to virus isolation. NCP BVDV strains are detected, and CP strains confirmed as BVDV, by immunofluorescence or immunoperoxidase staining techniques or reverse transcriptase PCR (RT-PCR).
■ TABLE 32.12
Diagnostic Testing for Bovine Virus Diarrhea Virus (BVDV)
| Clinical Form | Diagnostic Test | Specimen | Comments |
| Acute-transient | Virus isolation | Serum, whole | Whole blood is preferred. |
| BVDV infection | PCR | blood or nasal swabs | Viremia may be too transient for successful virus detection. |
| Virus isolation PCR | Nasal swabs | This is useful for establishing role of BVDV in bovine respiratory disease complex. | |
| Serum virus neutralization | Serum | Pair sera 3 to 4 weeks apart. Serologic testing for both types 1 and 2 BVDV should be performed. | |
| Fluorescent antibody | Postmortem | Tissues of choice include ileum, mesenteric lymph nodes, | |
| immunohistochemistry virus isolation PCR | tissues | spleen, thymus (calves), lung (pneumonia). | |
| Infertility | Serum virus neutralization | Serum | Serum bank for at-risk animals may be needed as seroconversion is complete when reproductive problem is noticed, or titers of affected cattle should be compared with those of unaffected cattle. Serologic testing for both types 1 and 2 BVDV should be performed. |
| Abortion | Serum virus neutralization | Serum | Pair sera 3 to 4 weeks apart. Serologic testing for both types 1 and 2 BVDV should be performed. |
| Serum virus neutralization | Fetal fluid | Presence of antibodies or BVDV is suggestive but not definitive for BVDV-induced abortion. Serologic testing for both types 1 and 2 BVDV should be performed. | |
| Fluorescent antibody immunohistochemistry virus isolation PCR | Fetal tissues | Tissues of choice include ileum, mesenteric lymph nodes, spleen, thymus, liver, and lung. | |
| Persistent | Virus isolation (IPMA) | Serum or whole | Testing should be repeated in 2 to 4 wk to confirm |
| infection | Antigen capture ELISA PCR | blood | persistent infection. For virus isolation, buffy coats should be used in these studies for calves younger than 6 months of age. |
| Antigen capture ELISA immunohistochemistry PCR | Skin biopsy | Blood samples should be retested in 3 to 4 wk to confirm persistent infection. Pools of skin or blood may be tested initially with PCR. | |
| Mucosal disease | Virus isolation PCR Fluorescent antibody immunohistochemistry Virus isolation PCR | Serum or whole blood Postmortem tissues | Both CP and NCP BVDV must be isolated. |
| Herd screening | PCR | Bulk milk tank Skin biopsy | Milk test screens only lactating cows; 300 cows per milk pool can be tested. |
| Serum virus | Serum from | Unvaccinated cattle 6 to 12 months of age should be tested. | |
| neutralization | sentinel cattle | Serologic testing for both types 1 and 2 BVDV should be performed |
CP, Cytopathic; ELISA, enzyme-linked immunosorbent assay; IPMA, immunoperoxidase monolayer assay; NCP, noncytopathic; PCR, polymerase chain reaction.
BVDV ANTIGEN DETECTION. Direct antigen detection methods with blood and a variety of fresh, frozen, or fixed tissues have been described for BVDV. These methods include immunohistochemistry, antigen-capture ELISA (ACE), and immunofluorescence. Because of standardization of methods, ACE testing kits have largely replaced the use of immunohistochemistry in diagnostic laboratories. Viral antigens can be detected from BVDV infected tissue, whole blood, serum, milk, or nasal swab samples.94 These tests are advantageous in that they allow rapid and inexpensive detection in comparison to virus isolation. In addition, amplification of antigen is not needed, so these tests are ideally suited for the detection of animals with considerable amounts of antigen: namely, PI carriers.
ACE methods on skin samples have become widely used to detect BVDV antigen and are applied for the detection of cattle PI with BVDV95 Skin biopsy samples may be obtained easily and inexpensively. A major advantage of testing by ACE on skin samples is that sampling can be performed on young PI calves, which could test negative by virus isolation, microplate virus isolation, and other serum or buffy coat testing because of inhibition by acquired colostral antibodies.96 Both the immunohistochemistry and ACE tests are ideally suited for the detection of PI animals, and reports indicate these tests very rarely detect acute infection in cattle.,98, With immunohistochemistry testing, PI animals can be differentiated from acutely infected cattle on the basis of location of viral antigen in the skin.100 In cattle vaccinated with modified live-virus vaccines, immunohistochemistry or ACE methods do not yield positive results.98,101
REVERSE TRANSCRIPTION POLYMERASE CHAIN REACTION. Since sequences of BVDV isolates became available, molecular techniques have been developed for diagnosis of BVDV, and this method of BVDV diagnosis has gained widespread use as a routine diagnostic method.95 An additional advantage of RT-PCR for diagnostics is that sequencing information can be readily and economically obtained, thus determining the BVDV species involved in the animal infection or herd problem. In comparison with traditional virus detection methods, PCR offers rapid results with high sensitivity and specificity. Development of commercial kits, with rapid and simple viral RNA extraction techniques, has made molecular techniques ideal for detection of viral genomic nucleic acids from many types of samples, including serum, whole blood, tissues, and bulk tank milk. The RT-PCR is more sensitive than virus isolation.102 Because of the high sensitivity of RT-PCR, it is used for pooled testing, and pooling of whole blood, serum, or milk samples is an economical way to detect BVDV infection in herd screening assays. A U.S. Department of Agriculture (USDA)-licensed PCR testing protocol and kit are now available for the pooling of ear notch samples.
SEROLOGY. Detection of antibodies to BVDV is another method of demonstrating virus infection; however, it is often difficult to distinguish antibodies produced in response to a natural infection, after vaccination, or as a result of transfer of maternal antibodies from dam to offspring. Interpretation of serologic results is difficult without important information about the herd being tested, such as the vaccination history. Serologic testing may be used to assess vaccine efficacy and vaccine protocol compliance, as well as to determine whether BVDV exposure has occurred in the herd.94 The BVDV-specific antibodies can be divided into two groups: neutralizing and nonneutralizing antibodies. Although there are several applicable methods, serum utilization, more appropriately called virus neutralization, and ELISA have been most commonly used for serologic diagnosis of BVDV Because of numerous technical and logistical challenges, the virus neutralization test is less useful than the ELISA.103
Treatment and Prognosis
No specific treatment is available for animals showing clinical signs of BVDV infection. Owners should be informed that severely ill animals may have mucosal disease, which ordinarily is fatal. The goals of therapy for cattle suspected of having acute BVDV infection are supportive care and prevention of secondary bacterial infection. Broad-spectrum antimicrobial agents, fluids, electrolytes, and vitamins may be indicated.
Prevention and Control
Control of any infectious disease relies on eliminating the reservoirs of the pathogen and limiting transmission from infected individuals to susceptible animals. PI animals are the reservoirs, and eliminating the potential for the birth of PI calves is key to prevention and control. The goal of BVDV control is to ultimately reduce production losses associated with BVDV infection. Development and implementation of herd health programs that involve vaccination and biosecurity to limit exposure of pregnant cattle to PI cattle are important for success of control. Therefore for prevention and control of BVDV, three major actions should be instituted: (1) identification and elimination of PI animals, (2) enhancing immunity through vaccination, and (3) implementing strict biosecurity measures to prevent introduction of PI carrier animals into a population of susceptible cattle. Each of these three principles has been applied104; however, BVDV continues to be a major problem to the cattle industry worldwide. Some European countries have successfully eradicated BVDV,105 and voluntary BVDV control programs have been implemented in the United States. Control and eventual eradication of BVDV can be possible in defined areas, even those with large and dense cattle populations.
IDENTIFICATION AND ELIMINATION OF PERSISTENT INFECTION IN CATTLE. The primary source of BVDV is cattle PI with BVDV because PI animals have a high and persistent viremia and shed BVDV from nearly all secretions throughout their lifetime. PI animals ideally should by identified and removed from breeding herds. In beef cow-calf operations with a controlled breeding season, PI animals should be removed before entry of a bull into the breeding herd.106 In this situation, all calves, replacement heifers, bulls, and nonpregnant dams without calves should be tested for BVDV PI status. Because PI cows always produce PI calves, a negative test result for a calf indicates a negative PI status of the dam.107 For any calves that test positive, the dam must be tested for BVDV PI status. Most PI calves result from an acute infection in the dam, rather persistent infection in the dam. Dams with negative test results may reenter the breeding herd; they possess high natural immunity to BVDV. In beef herds with a controlled breeding season, the BVDV status of all animals can be determined if no animals are pregnant at the time of testing. If cattle are pregnant at the time of testing, those animals must be segregated, and their calves' test results must be negative before return to the breeding herd.
In dairy herds and beef herds without a controlled breeding season, young calves should be tested and removed as soon as possible to avoid possible transmission to the breeding herd. Screening young calves for PI status is best accomplished with ACE or RT-PCR methods on skin biopsy samples. The use of skin samples for testing young calves is advantageous over other methods in that it is relatively easy for laymen to collect the samples, samples may be taken from calves that possess circulating maternal antibodies, and a single positive test result usually indicates PI status. Because acutely infected animals can produce positive RT-PCR results and occasionally positive ACE results, valuable breeding stock should be retested 30 days later with virus isolation on blood samples or ACE or RT-PCR assays on blood or ear notches.99
Screening all individuals in a herd for BVDV is very costly, especially considering the low prevalence of PI animals among cattle herds. Instead of whole-herd testing, monitoring strategies may be initiated, and these include evaluation of production records, BVDV evaluation of aborted fetuses, use of sentinel animals, pooling strategies for RT-PCR testing, and BVDV testing on sick or deceased individuals within the herd.106 Monitoring breeding records, calf morbidity and mortality rates, and weaning proportions are considered the minimal level of surveillance, and these involve the least expense, but this level of surveillance is not very sensitive for evaluating the presence of persistent infection with BVDV within a herd.11,106 As an example of the difficulty in utilizing clinical suspicion as a reason to perform herd testing, BVDV was isolated from cattle in 53% of herds in which there was no suspicion of the infection,108 and animals PI with BVDV were not identified in 81% of herds where veterinarians suspected BVDV was present.11 On the other hand, identifying BVDV in sick or dead animals or in aborted fetuses provides the justification for further whole-herd testing for animals PI with BVDV.
Because of high sensitivity, RT-PCR assays with pooled samples have been developed and tested to screen populations of animals for BVDV persistent infection.102,109 Sample types that have been utilized in RT-PCR assays of pooled samples include serum, whole blood, bulk tank milk, and skin. With pooling whole blood samples, a single PI animal is detectable in pools of up to 250 negative samples. RT-PCR assays have also been developed and evaluated for testing pools of skin biopsy samples.109 Pooled sample testing by RT-PCR is a rapid and cost-effective method for screening populations of cattle for animals PI with BVDV Individuals within the positive pools may be tests subsequently with ACE, immunohistochemistry, virus isolation, or RT-PCR methods, depending on the class of animals being tested. When testing young beef calves, it is important to perform all testing before the breeding season to identify and eliminate animals PI with BVDV that pose a risk for bred cattle.
ENHANCING IMMUNITY THROUGH VACCINATION. Many vaccines are commercially available for BVDV, and the majority of these USDA-licensed vaccines contain BVDV in combination with other bovine respiratory and reproductive pathogens, including BoHV-1, parainfluenza type 3, bovine respiratory syncytial virus, Leptospira spp., Campylobacter spp., M. haemolytica, Histopbilus somni, and Pasteurella multocida. Because of antigenic diversity among BVDV 1 and BVDV 2 strains, modified live- virus and inactivated vaccines containing both species are now widely available. Advantages and disadvantages of BVDV modified live-virus vaccines and inactivated vaccines have been described.110 One disadvantage of inactivated BVDV vaccines is that two doses are required for the initial immunization. A major problem with using inactivated vaccines is the widespread lack of compliance among producers.111 Failure to give a booster during the primary series and failure to regularly administer annual boosters with inactivated vaccines are common.111
Vaccines are an important component to BVDV prevention and control, and their effectiveness has been to limit disease from BVDV infection. In discussing BVDV vaccination with owners and producers, it is important to discuss reasonable expectations after vaccination and to remember that disease and infection are not synonymous terms. Experimental and field data indicate vaccination with either inactivated or modified live BVDV vaccines are effective at reducing or obviating clinical disease.54,112 In addition, modified live-virus vaccines containing BVDV 1 strains are effective at limiting or preventing clinical disease when vaccinated animals are subsequently challenged with a virulent BVDV 2 strain.113,114 Protection from clinical disease is important for stocker/backgrounder and feedlot operations, and immunity to BVDV has been demonstrated to be protective against bovine respiratory disease complex.115 Preconditioning cattle by vaccination against BVDV before an expected exposure (commingling and shipping) reduces the effects of exposure of cattle to BVDV.
Protection against viremia is the true measure of BVDV vaccine efficacy. To be truly efficacious, vaccination against BVDV should protect against viremia to prevent dissemination of virus throughout the host, including preventing infection of target cells of the reproductive tract that result in fetal infection. Since 2005, the focus for vaccine efficacy has shifted from protection against clinical disease to protection against fetal disease or infection. Published studies indicate the protection against fetal infections after BVDV vaccination varies from 60 to 100%. This wide degree of variation depends on whether the vaccine is inactivated or modified live-virus, the timing of challenge, and the degree of homology between the vaccine strains and the challenge strains. Although both modified live-virus and inactivated BVDV vaccines have been demonstrated to provide fetal protection, modified live BVDV vaccines provide greater protection, according to a meta-analysis of clinical trials.116 Fetal protection studies have been performed to evaluate commercial vaccines containing only BVDV 1 strains117,118 and commercial vaccines containing both BVDV 1 and BVDV 2 strains.119,120 From published studies, it appears that protection is superior when animals are challenged with 114121
strains from the same genotype.114,121 An extremely important observation regarding these fetal protection studies is that although protection may not be 100% against all strains of BVDV, the level of protection is far superior to that observed when proper vaccination is not utilized, as evidenced by very high numbers of PI animals among unvaccinated cattle.116
BIOSECURITY. Although BVDV prevention and control starts with identification and elimination of PI animals, strict biosecurity is essential to prevent reintroduction of the virus. Strict biosecurity is difficult for most cow/calf and dairy operations to achieve and is even more difficult or impossible for most stocker/backgrounder and feedlot operations to achieve. Biosecurity systems with isolation and testing of all incoming cattle are necessary to ensure that BVDV does not enter the farm. All purchased cattle should be isolated and tested for persistent BVDV infection before entry into the herd. In addition, isolation of new additions for three weeks before entry into the resident breeding herd should prevent transmission of BVDV from acutely infected animals. Most lapses in herd biosecurity involve purchasing cattle that are PI with BVDV or purchasing pregnant cattle in which the BVDV status of the fetus is unknown. If pregnant cattle are purchased, these animals should be isolated and their offspring should be tested to ensure that they are free of BVDV Semen should be used only from bulls that have test results negative for BVDV infection.
For purebred herds marketing genetically valuable embryos and livestock, it is important to test embryo transplantation recipients to ensure that they are not PI with BVDV. Exposure of cattle to other cattle or small ruminants at exhibitions should be limited, and animals should be quarantined for 3 weeks before reentry into the breeding herd. Most biosecurity principles instituted for BVDV control have the added benefit of disease control with other pathogens of cattle.