Infertility Caused by Abnormalities of the Female Genital Organs
Abnormalities Caused by Problems With Sexual Differentiation
Sexual differentiation occurs in three stages, each stage dependent on the previous one:
1. Chromosomal (genotypic or genetic) sex is determined at fertilization in mammals by the type of sex chromosome contributed by the sperm (X or Y).
In mammals, females are XX and males are XY.2. Gonadal sex is regulated by the Sry (sex-determining region of the Y chromosome) gene, which produces a protein called the HY antigen, as well as other sex-linked and autosomal genes downstream of Sry. Activation of this pathway induces testis formation in the bipotential gonads. The X-linked Daxl gene suppresses testis formation.
3. Phenotypic sex is regulated by substances produced in the male testes to cause regression of the female tract and formation of the male tract or, in the absence of a testis, formation of the female tract.
Chromosomal sex is determined at fertilization. All normal mammalian oocytes contribute an X chromosome in addition to one of each representative autosomal maternal chromosome. Sperm cells contribute either an X or a Y sex chromosome in addition to one of each representative autosomal paternal chromosome. Abnormalities in chromosomal sex occur because of nondisjunctional errors during either mitosis or meiosis. Fig. 43.1 shows some examples of chromosomal sex anomalies.
Monosomy X (XO)
Monosomy X is also known as Turner syndrome. Owing to the lack of a Y chromosome and the consequent Sry gene, but the presence of the Dax1 gene on the present X chromosome, the phenotype is female. Monosomy X is the most commonly reported chromosomal abnormality in mares. Animals with this syndrome often have a history of poor performance and lack of or sporadic reproductive cyclicity. Ovaries are typically
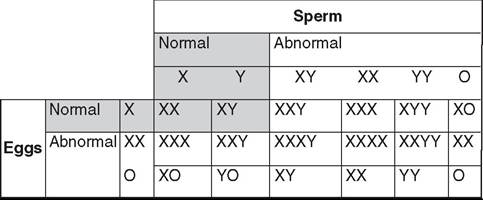
FIG.
43.1 Examples of chromosomal sex abnormalities.
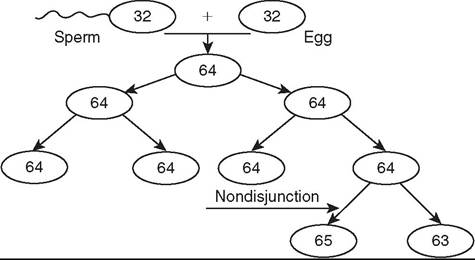
FIG. 43.2 Process of how a mosaic may occur. The haploid sperm and egg form a diploid zygote. Through the process of mitosis, diploid cells are replicated to always produce the same number of chromosomes. If, however, a chromosome pair does not separate during anaphase, called a nondisjunction event, cell lines that do not reflect a correct representation of the genome will be created and potentially propagated. If this happens with the sex chromosomes in the germ cell line, fertility of the individual or its offspring may be affected.
inactive, small, smooth, and firm. The uterus and cervix are usually hypoplastic. Externally the mare's genitalia may appear normal or underdeveloped.
XXY Syndrome
The genotype for XXY syndrome is part of Klinefelter syndrome. Because of the presence of a Y chromosome and the consequent Sry gene, affected individuals are phenotypically male, but probably owing to the presence of two copies of the Dax1 gene on the two X chromosomes, they generally have hypoplastic genitalia and reproductive organs. It is thought that some factor (Dax1 or some other) on the X chromosome must escape the inactivation process, which happens early in development (around day 7 or 8 in the horse). Testicular development and spermatogenesis are inhibited, resulting in small, flaccid testes and azoospermia. The testes may be retained or descended, but they are often small and soft. The penis may be normal or smaller than usual. Affected males often show normal libido and sexual behavior. Low testosterone concentrations may be noted. Infertility always accompanies this syndrome. Reports have been made in numerous species, including the horse.1,2
XXX
More is not better. A report of an infertile mare with the 65,XXX genotype confirms this.3 The mare had bilaterally small, inactive ovaries and a hypoplastic uterus and cervix.
MOSAICS. Mosaics are individuals that have at least two cell lines with different karyotypes arising from the same zygote (Fig. 43.2).
Phenotypes vary in accordance with the degree of mosaicism. Varying degrees of hermaphroditism and pseudohermaphroditism have been reported in many domestic species. Mosaics often have mixed gonadal dysgenesis, with an ovary and a testis, or ovotestes, owing to sex chromosome mosaic cell lines.4,5 These are true hermaphrodites.
Chimeras
Chimeras are individuals having cell lines from two different embryonic sources. This can occur experimentally or from the natural fusion of blastocysts in utero. The possibility has been reported from a suspected double ovulation and fertilization followed by blastomere fusion in the horse (64,XX/64,XY and 63,XO/64,XY genotypes reported). Freemartinism is a common occurrence in ruminants, resulting in chimeric twins.
Freemartins
Freemartinism is a phenomenon in ruminants in which an infertile female is twin to a male. The dizygotic occurrence happens when the blastodermic vesicles of the two zygotes fuse early in development (day 18 to 20 in cattle) and share embryonic tissue. The placentas fuse (day 30 to 50 in bovines), and they share blood throughout gestation. This occurs before gonadal differentiation at day 40 to 50. Both individuals are XX/XY chimeras. The Sry gene of the male twin causes the freemartin gonads to develop at least partially toward the male testis. The degree of differentiation varies with each freemartin, and many freemartin gonads remain undifferentiated. The shared circulation allows testosterone and AMH from the male twin, and possibly from the chimeric freemartin, to affect the freemartin genitalia, and so she lacks a cervix, uterus, uterine tubules, and cranial vagina. The vulva is fairly normal. The yearling freemartin fails to exhibit estrus, the udder and teats remain small, and the freemartin externally resembles a steer (only with a vulva).
Clinical diagnosis can be made by establishing a blind end to the vagina (no cranial vagina, no cervix) and confirmed with genotyping or use of polymerase chain reaction (PCR) to detect the Y chromosome in a phenotypic female. Among heifers born co-twin to a male, 92% will be freemartins.6The male twin may develop into a fertile adult, but these individuals show a higher incidence of infertility than bulls with a 60,XY genotype. Most male twins to freemartins become steers.
Freemartinism is less common in sheep than in cattle, but it does occur. It has also been reported in goats and pigs. With increased fecundity in modern animals, the authors observe the phenomenon more often than was reported in the past. This is because ovine freemartinism is rare with twins or triplets but much more common with quadruplets or quintuplets. A notable difference between ovine and bovine freemartinism is the marked masculinity of the ovine freemartin. Gonads within the inguinal canal resemble normal prepubertal testes, and those within the abdomen resemble cryptorchid testes from rams of normal XY gonadal sex. Many ovine freemartins also have epididymides, vasa deferentia, vesicular glands, and even cremaster muscles.
Gonadal Sex
Chromosomal sex determines gonadal sex. The Y chromosome has very few genes, and all of the ones studied play a role in sex differentiation. The gene located at the sex-determining region on the Y chromosome (Sry) has a DNA binding domain high-mobility group (HMG) box. It produces a protein called the HYantigen, and its action appears to be regulated by the transcription of other genes. Their actions initiate differentiation of bipotential embryonic gonadal tissue into testicular tissue. Other genes act downstream of Sry to support gonadal differentiation; these include Sox9, Gata4, Wt1, and Sf1, which act synergistically to promote testicular differentiation. Sox9 is a powerful promoter of testicular tissue differentiation. It is hypothesized that Sry upregulates Sox9.
In the absence of the Sry gene, the dual copies of the Daxl gene on the X chromosomes suppress the formation of testicular tissue by antagonizing the Sry gene and the synergy between Sfl and Wtl. Another gene, Wnt4, has been shown to support female development, and the absence of this gene in female mice results in masculinization.7 This evidence refutes the notion that female development is strictly a default process caused by the absence of Sry. Further research is elucidating other factors that actively promote the formation of ovarian tissue (e.g., Rspo1 β-catenin signaling). Factors affecting Sry, or any of the other genes downstream of Sry, or any factors governing female development will affect phenotypic sex.Abnormalities of Gonadal Sex: Sex Reversals
Sex reversals occur when the chromosomal sex and gonadal sex do not agree with each other. XX sex-reversed males and XY sex-reversed females are reported in many domestic species. These individuals are either XX males (testicular tissue), XY females (ovarian tissue), or true hermaphrodites (both ovarian and testicular tissue on separate gonads or the same gonad [ovotestes]). Sex reversals are relatively common in the horse and are considered to occur both sporadically and with familial inheritance.
XY SEX-REVERSED FEMALES (GONADAL DYSGENESIS). XY sex-reversed females have been reported to arise because of an absent or mutated (and nonfunctional) Sry gene. It is believed that the Sry chromosome is missing because of an abnormal meiotic exchange with the X chromosome. It is thought that in the sire, two crossing-over events occur between X and Y chromosomes during spermatogenesis. The offspring are subsequently infertile, and their external genitalia have a normal female appearance. The ovaries and uterus tend to be hypoplastic or even completely undifferentiated (“streak gonads”). These animals may be true hermaphrodites (ovotestes) or XY females (only ovarian tissue).
SRY-POSITIVE XX SEX-REVERSED MALES.
XX sex-reversed males conversely may arise from a translocation of the Sry gene onto the X chromosome. These animals may be true hermaphrodites or XX males (only testicular tissue). This condition has been reported in ruminants but not in horses.SRY-NEGATIVE XX SEX-REVERSED MALES. There are reports in horses of Sry-negative XX sex-reversed males.8,9 The exact mechanism of masculinization is still uncertain. Possibilities include Y-specific sequences other than Sry (which would require an XY individual with an inactivated or absent Sry gene), XX/XY chimerism within the testicular tissue, and a mutation in an autosomal or X-linked gene farther down the cascade of genes responsible for sex determination. XX sex-reversed males may have ambiguous sexual characteristics and may be true hermaphrodites or XX males (only testicular tissue).
Goats (especially Alpine, Saanen, and Toggenburg) present another classic, common example of Sry-negative XX sex- reversed males. Either the “polled” (hornless) gene is closely linked to an intersex locus or the polled gene itself is pleomorphic and controls both the hornless and intersex traits. This close linkage or pleomorphism is called the polled/intersex syndrome (PIS). A partial reason for the sex reversal is the deletion that affects a noncoding RNA (Pisrt1) and a transcription factor (FoxL2). The mechanism of testis induction has not been discovered yet. Elucidating this mechanism may be a big step in describing autosomal sex-determining factors in other species, including humans. The intersex gene is currently thought to work by mimicking the Sry gene and codes for the HY antigen.
The polled gene shows a dominant autosomal inheritance pattern, whereas the intersex trait shows an autosomal recessive inheritance pattern. A single dose of the P polled gene is enough to cause the polled trait, but a double dose is required to cause intersexuality in XX individuals. Therefore PP animals are hornless and infertile (if XX; fertile if XY); Pp individuals are hornless and fertile, and pp individuals are horned and fertile. Intersex individuals are always sterile and have a shortened vagina, large clitoris, bucklike head and neck, buck odor, and buck behavior. The gonads are testes or ovotestes and may be scrotal, inguinal, or abdominal. Even very masculinized intersexes with scrotal testes are azoospermic. A PP polled goat that has an XY genotype is usually initially fertile but often develops sperm granulomas later in life. The only way to avoid the intersex condition is to always breed a polled individual to a horned individual.
Phenotypic Sex
Gonadal sex determines phenotypic sex. Initially each embryo has both mullerian (paramesonephric) ducts and wolffian (mesonephric) ducts. Within testicular tissue, Sox9 triggers Sertoli cells to secrete mullerian-inhibiting substance (MIS; also known as anti-mullerian hormone [AMH]), which initiates the irreversible regression of the paramesonephric ducts. The action of MIS or AMH is further regulated by other genes and their proteins (Sf-1, Gata factors, Wt-1, Dax-1, and FSH). Wt-1 and Sf-1 synergistically enhance AMH transcriptional activity. Gata-4 enhances AMH promoter activity by directly binding to DNA and by synergistically interacting with Sf-1.
Leydig cells secrete testosterone that is converted by 5α-reductase to dihydrotestosterone. These two steroids promote the differentiation of male genitalia. Testosterone influences the differentiation of the wolffian ducts into the internal male genitalia (vasa deferentia and epididymides), whereas dihydrotestosterone stimulates the formation of the seminal vesicles and male urethra from the urogenital sinus, and the penis from the genital tubercle. In the absence of these testicular hormones, the wolffian ducts regress and the mullerian ducts become the female internal genitalia.
In the presence of two X chromosomes, the double dose of Dax-1 has an inhibitory effect on the synergistic relationships between Sf-1 and Wt-1 and Gata-4, preventing their support of AMH production.10 The primitive sex cords (gonadal cords) degenerate in the medulla and remain in the cortex (opposite in the horse). Subsequently there is no communication between the gonad and the mesonephros. In the absence of AMH, the mullerian ducts persist as the oviducts and fuse to form the uterus and cranial vagina. In the absence of testosterone, the wolffian ducts regress. Vestigial traces of these are located in the mesentery of the ovary, the epoophoron, the paroophoron, and Gartner's ducts. The tissues that form the round ligament of the uterus are analogous to the male gubernaculum.
Abnormalities of Phenotypic Sex
Abnormalities of phenotypic sex occur when the chromosomal and gonadal sex agree (XX with ovaries or XY with testes) but the external and/or internal genitalia do not correlate or are ambiguous. Affected animals are the male and female pseudohermaphrodites. The condition can occur because of insensitivity of androgen receptors or because of any abnormality along the pathway that may affect the intervening hormones, such as the conversion of 5α-reductase to dihydrotestosterone.
TESTICULAR FEMINIZATION. Testicular feminization is reported in domestic species, including the horse.11 Patients have external genitalia that are either female or ambiguous in appearance. The vagina may be blind-ending, or the uterus may be hypoplastic. The gonads are testicles, although they are usually abdominal or inguinal. Male behavior may be reported in horses. The problem lies in the gene for the androgen receptor, located on the X chromosome received from the dam. This X-linked recessive inheritance has been demonstrated in both humans and horses. Affected individuals are male pseudohermaphrodites. The condition has been diagnosed in multiple horse breeds. Serum testosterone concentration is often increased because of loss of negative feedback; a subnormal testosterone response to an hCG test is expected.