Physiology
The major functions of the large intestine are extraction of water and electrolytes from the ileal effluent, storage of feces, and defecation. In addition, microbial fermentation of organic matter that escapes digestion and absorption in the small intestine also occurs in the large intestine.
Absorptive processes and microbial metabolism occur in the proximal colon whereas fecal storage and elimination occur in the distal colon. These functional differences may be attributed to regional differences in patterns of colonic motility.6.3.1 Motility
The majority of muscular contractions arising in the proximal colon are retrograde peristaltic contractions, which are initiated in the transverse colon and are propagated towards the cecum.2 Termed antiperistalsis, this type of motility slows the transit of colonic contents and enhances mucosal absorption of fluid and electrolytes. The transverse colon plays the most important role in the proximal colon for mixing, storage, and dehydration of fecal contents.
Throughout the colon, rhythmic segmentation, originating in the circular muscle layer, moves the contents short distances in both antegrade and retrograde directions, preventing rapid transit. This promotes absorption of the remaining water and electrolytes. Coordinated peristalsis can be observed in any part of the colon, but is the predominant pattern of motility in the mid-portion of the colon. Peristaltic waves are tonic rings of constriction, transmitted by the longitudinal muscle layer, which serve to move colonic contents aborally.
Spontaneous giant migrating contractions, or mass movements, are the predominant form of motility in the distal large intestine. These powerful smooth muscle contractions originate in the proximal colon and migrate in an aborad direction over a segment or the entire length of the colon, moving colonic contents towards the rectum in preparation for defeca- tion.5
Intrinsic smooth muscle properties, intrinsic neurons, extrinsic nerves, and neuroendocrine polypeptides influence colonic contractile activity.
Control of peristalsis and rhythmic segmentation in the colon is determined by slow wave activity, which is an inherent myoelectric property of smooth muscle. Slow waves are generated by ion fluxes across the smooth muscle cells. The frequency of slow wave activity determines the rate of contraction, while the number of spike potentials superimposed upon slow waves and generated during the depolarization phase determines the strength of contractions. Slow waves are less frequent in the proximal colon than in the distal colon, which helps to impede transit and promotes the extraction of water and electrolytes. In contrast to the small bowel, slow waves in the large bowel are generated in the circular muscle layer. There are multiple pacemakers in the colon that propagate slow waves over a short distance. However, a single pacemaker exists in the transverse colon, from which slow waves are spread most often in an orad direction and are associated with antiperistalsis. Giant migrating contractions result from prolonged bursts of electrical activity, which span several slow wave cycles and appear to be independent of slow wave activity. Differences in the intrinsic mechanical properties of colonic longitudinal and circular smooth muscle have been demonstrated in cats and contribute to regional specialization of motility. This is most notable in the proximal colon where antiperistaltic activity of the longitudinal smooth muscle layer predominates over the segmental activity of the circular muscle layer.The intrinsic nervous system is essential for normal colonic motility and can function independently of the extrinsic nervous system. Reflexes mediated through efferent cholinergic neurons stimulate segmental contractions and peristalsis. Stimulation of mechanoreceptors in the muscular layers that respond to distension, or of chemoreceptors within the mucosa that respond to luminal contents result in muscular contractions.
In addition to acetylcholine and norepinephrine, a variety of other neurotransmitters have been identified that are released from the neurons of the intrinsic nervous system.
Many of these are peptides and include neurotensin, cholecystokinin, substance P, somatostatin, and 5-hydroxytryptamine.6,7 The response of colonic smooth muscle to neurotransmitters varies throughout the colon. Regional differences have been observed in smooth muscle responses to neurotensin and cholecystokinin, both of which are more potent stimuli of proximal than distal colonic smooth muscle activity.8 These differences may, in part, explain the different motility patterns and functions in the proximal and distal colon.Extrinsic neural and hormonal regulation of colonic motility is poorly understood. In general, extrinsic parasympathetic activity stimulates and sympathetic activity inhibits segmental contractions. The major function of the extrinsic nervous system is in the distal colon, where it participates in the defecation reflex.
The defecation reflex may be initiated in response to ingestion of a meal (gastrocolic reflex) as well as luminal distension in the distal colon and rectum. Increased tension in the rectal wall stimulates intramural receptors that transmit impulses along afferent parasympathetic pathways within the pelvic nerve to the sacral spinal cord. Efferent nerve fibers contained within the pudendal, hypogastric, and pelvic nerves complete the reflex arc. Stimulation of these fibers initiates a motor response that causes contraction of colonic and rectal smooth muscle, and relaxation of the internal and external anal sphincters, allowing evacuation of the rectum and distal colon. Defecation is usually preceded by giant migrating contractions of the distal colon.5 The defecation reflex can be inhibited by voluntary contraction of the external anal sphincter. Receptive relaxation of the distal colon and rectum accommodates fecal storage until the next defecation reflex is initiated.
6.3.2 Water and electrolyte transport
The large intestine normally absorbs a smaller quantity of water than the small intestine, but does so much more efficiently.
Approximately 90% of the water entering the large bowel is absorbed.2,9 Water absorption in the large bowel occurs passively across an osmotic gradient principally created by the absorption of sodium. The large bowel has a maximum absorptive capacity for water. When fluid flow from the small intestine exceeds this absorptive capacity, when there is excessive colonic fluid secretion, or when colonic absorption is decreased, diarrhea ensues. Therefore, the capacity of the large bowel to absorb water principally determines whether or not diarrhea is present with either small or large bowel diseases.Sodium absorption in the proximal colon appears to occur principally via chloride-coupled sodium transport. This process results from two separate ion transport mechanisms working in concert to facilitate the movement of sodium and chloride across the apical membrane of the epithelial cells. An even exchange of sodium with potassium and chloride with bicarbonate maintains electrical neutrality.
The large bowel secretes potassium by way of an active conductance mechanism at the apical membrane of the colono- cytes. In contrast, active potassium absorption occurs in the distal colon through a K+∕H+ exchange.
Mineralocorticoids and glucocorticoids stimulate sodium absorption and potassium secretion in the distal colon. These substances affect colonic transport by increasing the permeability of the apical membrane to sodium and potassium, and by enhancing the activity of the Na+∕K+ ATPase pump. Colonic epithelial cells also secrete bicarbonate. Bicarbonate neutralizes the acid produced by colonic bacteria, which can be irritating to the mucosa. The secretion of bicarbonate is cou-
pled to the absorption of equal numbers of chloride ions and may occur independently of sodium absorption.
6.3.2 Mucus secretion
Mucus is the principal secretory product of the large bowel. Mucus acts as a lubricant and facilitates the passage of large bowel contents and protects the mucosa from mechanical or chemical injury.
Various agents (microorganisms, enterotoxins) can be bound by mucus, thus preventing their attachment, entry into, or their adverse effects upon the colonic mucosa.9,10 Direct tactile stimulation of the mucus cells by luminal contents and intramural reflexes primarily regulate the production of mucus. The large bowel is capable of secreting large volumes of mucus, water, and electrolytes when inflamed. This response serves to dilute the irritating factor and promote colonic distension and evacuation of colonic contents. Marked increases in mucus secretion can also be caused by stimulation of parasympathetic activity.6.3.3 Colonic microflora
The large bowel contains the highest concentration of bacteria within the GI tract, with a single gram of feces containing up to 1011 organisms.2 Nearly 50% of the dry weight of feces consists of bacteria. Anaerobic (spore- and non-spore-forming) bacteria predominate, accounting for up to 90% of the large intestinal microflora. Bifidobacterium spp. and Bacteroides spp. are found in the highest numbers, with lesser numbers of Clostridia spp. The predominant aerobic bacteria found in the large bowel are Lactobacillus spp., Enterobacteriacea spp., and Streptococcus spp.
The physiological mechanisms that are important to maintain a normal colonic microflora and to prevent disease induced by bacterial overgrowth or colonization with pathogenic organisms include normal colonic motility, maintenance of the mucosal barrier, and local immune factors. Interactions of resident microflora, diet, and orally administered or enterohepatically circulated antibacterial agents also influence the colonic microflora.
The normal large intestinal microflora resist colonization by other, potentially pathogenic, organisms by producing metabolic products that inhibit growth of other bacteria and by competing for mucosal attachment sites and nutrients. The use of antibiotics with an anaerobic or broad spectrum may upset this sensitive balance and may result in an overpopulation of potential pathogens.
Colonic bacteria metabolize carbohydrates, proteins, and lipids. Carbohydrates are fermented into acidic short-chain fatty acids (acetate, propionate, and butyrate) and gases (hydrogen, methane, and carbon dioxide). Luminal bicarbonate neutralizes most of the acids, resulting in the production of carbon dioxide and water. Luminal fatty acids may inhibit replication of pathogenic bacteria. Absorbed fatty acids are either metabolized by the colonic epithelium (butyrate) or are transported to other tissues and used as an energy source (acetate, propionate, or butyrate).
6.3.4 Immune function
The immune functions of the intestinal tract are extremely complex and our knowledge of them is constantly expand- ing.2,11 The major functions of intestinal immunity include: tolerance to orally administered antigens, locally protective responses, and systemic and mucosal dissemination of stimulated B- and T-lymphocytes.12 Natural intestinal defense mechanisms include both mechanical and immunological factors, which act in concert to protect the host.1 These mechanisms are particularly important because of ingestion and digestion of nutrients, ingestion of potentially pathogenic organisms and foreign substances, and the extensive resident bacterial flora contained within the intestines. Important mechanical factors include the mucosal epithelial barrier, intestinal motility, mucus production, and the normal resident bacterial flora.
The gut-associated lymphoid tissue (GALT) is compartmentalized into afferent and efferent sites.12 The afferent arm of the GALT consists of mucosal lymphoid follicles, Peyer’s patches, and the mesenteric lymph nodes. Effector functions are performed in the lamina propria and intestinal epithelium.10,12 The function of the lymphoid follicles is to capture and process antigens in order to initiate an appropriate immunological response. The GALT contains both B- and T-lymphocytes and antigen-processing cells, such as macrophages and dendritic cells. The germinal centers of the lymphoid follicles contain IgA-producing B-lymphocytes. Specialized epithelial cells, termed membranous or M-cells overlie the lymphoid follicles. These cells trap soluble or particulate antigens and whole microorganisms and channel them towards the antigen-processing cells.11
Intraepithelial lymphocytes (IEL) are principally suppressor T-cells, many of which are CD8+.12 Mast cell progenitors are also found in this compartment. In the mucosal lamina propria, IgA-producing B-cells predominate the B-cell subtype, while the majority of the T-cells are of the helper subset, many of which being CD4+.12 Plasma cells, mast cells, macrophages, and eosinophils are also present in the lamina propria. Besides mediating immediate hypersensitivity, mast cells also may play a role in delayed hypersensitivity, inflammation, cytotoxicity, and immunoregulation.
Plasma cells within the mucosa produce IgA, which is bound to a secretory component on the epithelial cell membrane and released into the gut lumen. Secretory IgA prevents the attachment and uptake of antigens by the intestinal mucosa by binding them. IgA in combination with protective mechanical factors serve to exclude antigenic material from the mucosa.
Antigens captured by M-cells or those penetrating the mucosal epithelium initiate an immune response. Following antigen processing, the immune system undergoes regulated expansion to produce clones responsible for cell-mediated immunity, humoral immunity, immune memory, and immunoregulation. The immune response results in sensitivity (elimination) of or tolerance to the antigen.
Cell-mediated immunity involves cellular cytotoxicity and synthesis of cytokines. When stimulated, T-helper-1 lymphocytes secrete IL-2, IFN-γ, and TNF-α, which support the cell-mediated immune response.12 Lymphocytes in the epithelial and mucosal compartments possess cytotoxic potential. Macrophages also may become cytotoxic. T-helper-2 lymphocytes secrete IL-4, IL-5, and IL-10, which support antibody production.12
Following antigen penetration of the mucosal barrier, nonIgA immunoglobulins become involved in the humoral immune response. IgM, IgG, and IgE participate in host defense by the processes of opsonization, complement fixation, and promotion of an inflammatory response, all properties that IgA lacks. IgE is also important in the mediation of immediate hypersensitivity. When antigen binds to IgE molecules fixed to mast cells, degranulation of the mast cells occurs, releasing inflammatory mediators (e.g., histamine, serotonin, leukotrienes, and others). This release of inflammatory mediators by mast cells may be important in the expulsion of parasites.
Immunological tolerance is an important feature of the GI immune system. Tolerance results from the induction of IgA, T-cell deletion, anergy, and immunosuppression.11 Most intraluminal antigens processed via M-cells induce tolerance. Both IL-10 and TGF-β are down-regulatory cytokines that help to promote tolerance.12 However, potentially harmful antigens that penetrate the mucosal barrier stimulate lymphocytes producing IgM, IgE, IgG, and proinflammatory cytokines, such as IFN-γ, IL-12, and IL-6.11 Tolerance is necessary to prevent mucosal and systemic injury as a result of a continuous inflammatory response to benign, persistent intraluminal antigens. A breakdown of this tolerance response may be involved in the pathogenesis of inflammatory bowel disease (IBD).
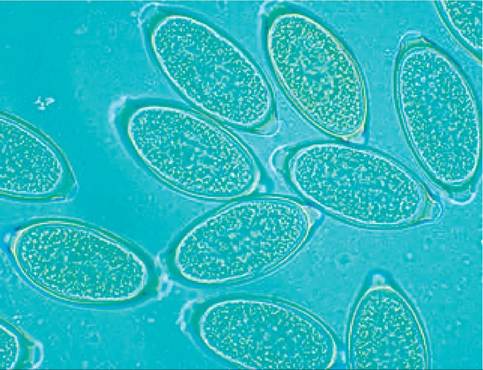
Figure 6.3:
Whipworm eggs. Close up of a group of whipworm eggs in a fecal flotation.
6.4