Physiology
The exocrine pancreas has several important functions. Most importantly, the acinar cells, which make up the bulk of the exocrine pancreas, synthesize and secrete many digestive enzymes and zymogens of digestive enzymes (Table 8.1).2 These digestive enzymes produced by the pancreas are required for digestion of the diet.
While the digestive tract is characterized by redundancy (i.e., the digestion of each dietary component is facilitated by more than one process), most dogs and cats require at least some pancreatic function for digestion. However, there are isolated cases of patients that have virtually no residual exocrine pancreatic function and which do not have any clinical signs of maldigestion. In addition to the synthesis and secretion of pancreatic digestive zymogens and enzymes, the exocrine pancreas also synthesizes and secretes a variety of other molecules (Table 8.1), such as intrinsic factor, which is crucial for cobalamin absorption; colipase, which is needed to reverse the inhibition of pancreatic lipase by bile salts; a trypsin inhibitor; antibacterial factors; and probably certain trophic factors that have been hypothesized to affect intestinal mucosal thickness.2,3Some digestive enzymes are synthesized and secreted as active enzymes, while others are synthesized and secreted as inactive proforms or zymogens.2 In general, any enzyme that is capable of digesting components of the cellular membrane, such as proteins and phospholipids are secreted as zymogens, while those enzymes that digest components that are normally located inside of organelles or the nucleus are secreted as active enzymes. Examples of enzymes that are secreted as zymogens (Table 8.1) are trypsin (i.e., trypsinogen), chymotrypsin (i.e., chymotrypsinogen), elastase (i.e., proelastase), and phospholipase (i.e., prophospholipase).2 In contrast, examples of enzymes that are secreted as active enzymes (Table 8.1) are lipase, amylase, desoxyribonuclease, or ribunuclease.2
Pancreatic enzymes and zymogens of pancreatic enzymes are synthesized by protein synthesis; that is, DNA is transcribed to mRNA in the nucleus of the acinar cells, and the mRNA in turn is read at the ribosomes to be translated into a polypeptide strand, the pre-pro-enzyme or pre-enzyme (in the case of enzymes that are secreted as active enzymes).
The pre-enzymes and pre-pro-enzymes are then internalized into the rough endoplasmic reticulum. During the process of internalization, a small signal peptide is removed, leaving the pro-enzyme or enzyme. The zymogens and enzymes are then processed by the Golgi apparatus, where they undergo glycosylation and other post-translational changes. At the distal end of the Golgi apparatus, the zymogens and enzymes are then packaged into zymogen granules. These zymogen granules are then released into the duct lumen by exocytosis.Peptides, amino acids, and fatty acids containing eight or more carbon atoms or their monoglycerides are the most important stimuli of CCK release from the neuroendocrine cells in the duodenum and jejunum. CCK stimulates gall bladder contraction and also leads to the secretion of zymogen granules from the acinar cells into the pancreatic duct system. A small amount of zymogen granules are also released into the vascular space.
Activation of pancreatic trypsinogen to trypsin is by hydrolysis, which is catalyzed by enteropeptidase, another serine protease that is secreted by the duodenal mucosa. In turn, activated trypsin then activates more trypsinogen molecules and other zymogens.2 Pancreatic digestive enzymes are crucial for the digestion of most dietary components, but oral, gastric, and brush border enzymes also contribute to this process. This contribution can be quite significant; for example, under physiological conditions, a significant portion of dietary fat is digested by gastric lipase.4
As said before, the main function of the exocrine pancreas is to digest dietary components. One important dietary component in carnivores and omnivores is meat, which includes pancreatic tissue; thus, the pancreas is always in danger of digesting itself. However, there are several mechanisms in place to prevent such autodigestion.5 First of all, the pancreas synthesizes and secretes all the enzymes that can be dangerous to itself as inactive preforms or zymogens.
Secondly, these zymogens are stored in zymogen granules within the acinar cells and are kept strictly separate from the lysosomes as lysosomal enzymes are capable of activating zymogens and so such strict separation is important to prevent pancreatic autodigestion.5 Thirdly, pancreatic zymogens are inhibited from autoactivation by the adverse conditions (e. g., pH) within the zymogen granules. This may not completely abolish autoactivation, but certainly decreases its likelihood. Fourthly, there is a small inhibitory molecule, pancreatic secretory trypsin inhibitor (PSTI), that is co-synthesized, co-transported, and co-stored with the pancreatic zymogens. PSTI inhibits any prematurely activated trypsin molecules, thereby preventing the cascade of pancreatic enzyme activation. PSTI probably plays a crucial role in preventing pancreatitis in normal individuals. In humans, hereditary pancreatitis is caused by mutations of the genes encoding PSTI (i.e., SPINK gene) or trypsinogen.6 These mutations either lead to the production of non-functional PSTI or of a trypsin that is not effectively inhibited by PSTI. Recently, mutations have also been identified in the SPINK gene of Miniature Schnauzers with pancreatitis.7 Fifthly, there is a unidirectional flow of pancreatic juice in the pancreatic duct. Pancreatic zymogens are activated in the small intestine and it is crucial that these activated enzymes cannot return to the pancreas where they could lead to pancreatic autodigestion and pancreatitis. Sixthly, and finally, if all of these safety mechanisms are overcome and activated pancreatic enzymes escape into the vascular space, they are quite effectively removed by proteinase inhibitors, such as α1-proteinase inhibitor (α1-PI) and α2-macroglobulin.8 Trypsin bound to α1-PI is quickly transferred to α2-macroglobulin, which is then trapped in the reticuloendothelial system of the spleen and thus removed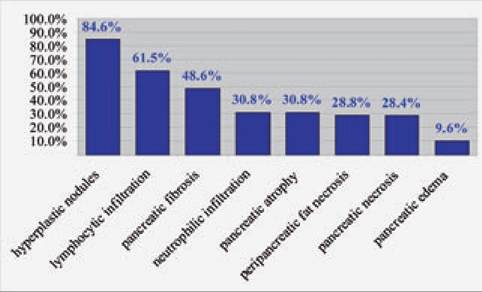
Figure 8.3:
Frequency of histopathological lesions in dogs.
This figure shows the frequencies of histopathological lesions of the pancreas of 73 dogs presented for necropsy to the Department of Pathology at the Animal Medical Center in New York. The dogs evaluated were affected by a wide variety of diseases and died of various causes. Each pancreas was sectioned every 2 cm and each section was evaluated for neutrophilic infiltration, lymphocytic infiltration, pancreatic necrosis, peripan- creatic fat necrosis, edema, pancreatic fibrosis, pancreatic atrophy, and pancreatic hyperplastic nodules. With the exception of hyperplastic nodules, all the other lesions can be considered as evidence of previous or current pancreatic inflammation.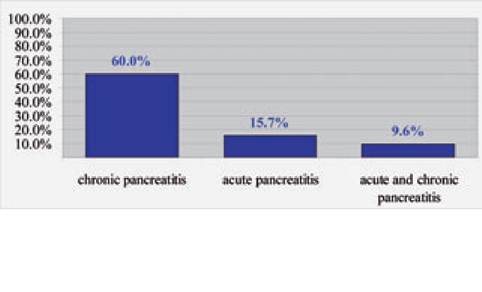
Figure 8.4:
Frequency of histopathological lesions in cats. This figure shows the frequencies of histopathological lesions of the pancreas of 115 cats presented for necropsy to the Department of Pathology at the University of California, Davis. The cats were affected by a wide variety of diseases and died of various causes. Three biopsies were taken from each pancreas (left limb, right limb, and pancreatic head) and each section was evaluated separately for evidence of either acute or chronic pancreatitis. Lesions that were considered to be suggestive of acute pancreatitis were interstitial edema and /or necrosis of mesenteric fat or neutrophilic infiltration of the pancreas. Lesions that were considered to be suggestive of chronic pancreatitis were lymphocytic infiltration, interstitial fibrosis, and cystic acinar de- generation.13
from the circulation. There may be other protective mechanisms that are as yet unknown, but it is still quite impressive how many protective mechanisms are in place to prevent the pancreas from damaging itself.
8.3