Specific Diseases ofthe Musculoskeletal System
Viral Diseases
Caprine Arthritis Encephalitis
First recognized in the early 1970s, CAE has become established as a significant and costly disease of goats. Goats exposed to the causative lentivirus may remain subclini- cally infected or develop one or more of several clinical syndromes, which include arthritis, progressive paresis or other neurologic dysfunction, an induration of the udder with hypogalactia, chronic interstitial pneumonia, or progressive weight loss.
Arthritis is the most common clinical presentation and is discussed here.Etiology
The CAE virus is an enveloped, positive, single-stranded RNA virus in the lentivirus genus of the family Retroviridae. Other lentiviruses, or slow viruses, include the maedi-visna (MV) virus of sheep (also known as the ovine progressive pneumonia or OPP virus), the equine infectious anemia virus, the bovine immunodeficiency virus, the feline immunodeficiency virus, the simian immunodeficiency virus, and types 1 and 2 of the human immunodeficiency virus (HIV) associated with acquired immunodeficiency syndrome (AIDS). Historically, CAE and MV viruses were considered to be similar but distinct viruses with strong host species predilection for goats and sheep, respectively. However, recent phylogenetic studies indicate that these distinctions do not hold strongly, that genetic recombination occurs, and that there is evidence for cross-species transmission. The two viruses are now classified together in the small ruminant lentivirus (SRLV) serogroup, and within this serogroup there are four separate genotypes containing various subtypes. The CAE virus is classified in SRLV genotype B and MV virus in the SRLV genotype A, with the potential for both genotypes to infect both goats and sheep. In other words, the SRLV can be considered as host adapted, but not strictly host associated. The phylogenetic basis for this revision and its diagnostic and epidemiologic significance are discussed in more detail in the CAE-MV section of Chapter 5.
These are cell-associated viruses characterized by the presence of a magnesium-dependent, RNA-dependent DNA polymerase, or reverse transcriptase, which permits the production of proviral DNA from the viral RNA genome using the host cell machinery (Cheevers and McGuire 1988). Typical of lentiviruses in general, CAE virus infection is considered to be lifelong, with the lentivirus genome integrating into the host DNA, and serum antibodies to the virus, if not of colostral origin, are considered synonymous with infection.
The CAE viral genome consists of three essential retroviral genes - gag, pol, and env - as well as the regulatory/ accessory genes - tat, rev, and vif. The genome produces five structural proteins and four non-structural proteins. The env gene encodes for the envelope glycoprotein gp135 or surface (SU) protein and the transmembrane protein. Antigenic variation in the SU protein occurs as a result of env mutation and is responsible for the biologic and serologic variation of CAE virus isolates (de la Concha- Bermejillo 2003). The pol gene encodes for the viral enzymes reverse transcriptase, protease, endonuclease/ integrase, and dUTPase, involved in transcription and protein synthesis during viral replication. The molecular biology of CAE virus and other human and animal lentiviruses has been reviewed (Clements and Zink 1996; Bose et al. 2015).
CAE virus is susceptible to heat, detergents, and formaldehyde.
Epidemiology
The neurologic form of CAE was the first to be described in the United States, in 1974 (Cork et al. 1974). The causative virus was first isolated from a case of arthritis in 1980 and the relationship of the two forms of the disease was recognized at that time (Crawford et al. 1980). The initial name of the disease, viral leukoencephalomyelitis of goats, was gradually replaced by the currently applied name, CAE.
A global serologic survey reported in 1984 provided much initial information about the distribution of CAE virus in goats (Adams et al.
1984). The highest prevalence occurred in countries with long-established, intensive goat dairying industries, namely Canada, France, Norway, Switzerland, and the United States. The seroprevalence exceeded 65% in all of these countries. In countries actively importing dairy goats, such as Kenya, Mexico, New Zealand, and Peru, the overall prevalence of CAE infection was often less than 10%. However, it was highest in imported dairy goats, next highest in local goats in contact with imported goats, and least prevalent or non-existent in goats not having contact with imported goats. Indigenous African goat breeds tested in Somalia, Sudan, and South Africa were free of infection. The same pattern of prevalence in imported, contact, and indigenous goats was reported from Jamaica (Grant et al. 1988) and from Algeria (Achour et al. 1994).In 24 states of the United States, 81% of 1160 goats tested were positive for CAE virus infection (Crawford and Adams 1981), while a subsequent, larger survey in 28 states reported only 31% of individual goats positive, but 73% of herds positive (Cutlip et al. 1992). In Great Britain, 10.3% of herds tested and 4.3% of all goats tested were positive (Dawson and Wilesmith 1985). In New South Wales, Australia, 82% of 115 herds were infected. Among the five different dairy breeds tested, the prevalence varied from 26.8% to 43.4%, while the prevalence in Angora goats was only 4.8% (Grewal et al. 1986). In South Australia, prevalence in dairy goats tested over three years ranged from 18% to 45%, while the combined prevalence in Angora, Cashmere, crossbred, and feral goats was 0.1% (Surman et al. 1987). In most cases, Angora goats with serologic or clinical evidence of CAE infection have a history of contact with infected dairy goats. A New Zealand survey indicated a prevalence of 1.5%, with a distribution pattern associated with goats imported from Australia (MacDiarmid 1983).
Additional reports on the prevalence of CAE in different countries and regions continued to appear.
There are multiple reports from Europe. In Norway, 51 dairy goat herds distributed throughout the country were tested and 42% of animals and 86% of herds were positive (Nord et al. 1998). In Sweden, approximately 90% of the dairy goat herds, comprising mostly the endangered native Swedish Landrace breed, were found to be infected with CAE (Lindqvist 1999). In Spain, where the disease was suspected but not yet confirmed, a survey of 22 dairy herds identified 77.3% of the herds and 12.1% of the individual goats tested as positive (Contreras et al. 1998). In Hungary, where 30% of goats tested were positive, the prevalence of infection was associated with purchase of breeding stock from farms that had imported purebred milking goats and an association was also made with large herd size (Kukovics et al. 2003). In a study from South Tyrol in Italy, 38% of herds and 23.6% of goats tested were positive, with goats older than 26 months having a significantly higher prevalence than younger goats. Goats born indoors had a significantly higher prevalence than those born outside, and dairy goats in commercial herds had a higher prevalence than dwarf goats kept as pets (Gufler et al. 2007).In Latin America, CAE is present in Brazil and, as in other places, it is more common in imported or crossbred dairy goats than indigenous non-dairy breeds. One survey of dairy goat herds in Rio Grande Norte state revealed 57% of the herds with at least one positive goat (de Souza e Silva et al. 2004). In Yucatan state of Mexico, seroprevalence was very low, but all the animals identified as positive were exotic-breed dairy goats that had been imported into the Yucatan (Torres-Acosta et al. 2003).
From the Middle East, a Jordanian survey found that 23.2% of herds and 8.9% of goats sampled were positive for CAE (Al-Qudah et al. 2006). In Asia, Japan was presumed to be free of CAE, but a serosurvey reported in 2006 indicated a seroprevalence of 21.9% in goats from around the country as well as clinical disease in native Japanese Shiba goats, including arthritis, mastitis, and pneumonia (Konishi et al.
2006). Newer reports from Africa are limited. South Africa is reported to be “relatively free” of CAE and maintains strict import regulations to minimize risk, while Kenya is reported to have eradicated the disease (Werling and Langhans 2004).Transmission
CAE virus can be transmitted vertically and horizontally. The main route of vertical transmission is from dam to kid via ingestion of virus-contaminated colostrum and milk. The main route of horizontal transmission is by direct or close contact with infected animals. Sexual transmission, though possible, plays a limited role in the spread of CAE. Risk factors associated with the transmission of CAE have been reviewed (Rowe and East 1997; Blacklaws et al. 2004). The strong association of CAE virus infection with intensively managed dairy goats is due largely to management practices that facilitate transmission of the virus from does to kids in the perinatal period as well as horizontally between adults (East et al. 1987, 1993; Greenwood et al. 1995).
In the perinatal period, the major mode of transmission of CAE virus to kids is by direct suckling of their own infected dam or by consuming pooled milk or colostrum from other infected does. Virus may be present in colostrum and milk as free virus or incorporated into somatic cells. Even subclinically infected does can shed virus into colostrum and milk. Because common methods of identifying infected goats, such as the agar gel immunodiffusion (AGID) test, can produce false-negative test results, all colostrum and milk derived from does in known infected herds is potentially suspect as a source of virus unless heat treated or pasteurized, as discussed later.
Additional potential sources for infection of kids in the perinatal period include in utero infection, exposure to virus in birth fluids during kidding, licking of the kid by the doe after birth, or possibly aerosol transmission between a coughing doe and her kid. Experimental evidence for the presence of CAE virus in utero or in saliva, birth fluids, or aerosols is limited compared to what is known about the virus in milk and colostrum.
However, epidemiologic observations with CAE in goats as well as experimental evidence of the transmission of the MV virus from ewes to lambs suggest that these routes of transmission can occur and that CAE control programs should take them into account in order to succeed (Rowe and East 1997; Blacklaws et al. 2004).To date, there are still no definitively documented cases of in utero infection of kids with CAE, although there are several indications that this is a possible route of infection. Some cesarean-derived or vaginally delivered kids raised in isolation have seroconverted in the first few months of life (Adams et al. 1983; East et al. 1993). There is also a report of uterine lesions in a goat with the arthritic and pneumonic forms of CAE infection (Ali 1987). More recently, using polymerase chain reaction (PCR) techniques, CAE virus-infected epithelial cells were detected in the uterus and oviducts of CAE virus-infected goats (Fieni et al. 2003; Ali al Ahmad et al. 2012).
Horizontal transmission also plays a significant role in the spread of CAE. An early indication of this related to international trade in livestock, when it was observed that local goats became infected, seroconverted, or even manifested clinical disease when imported, infected purebred dairy goats were placed in their midst in countries not previously recording the disease. Another strong indication of horizontal spread is the common finding in many studies that seroprevalence increases with age in infected herds. Some seroconversion in goats less than 1 year of age may result from neonatal infections with delayed humoral response. However, it is highly unlikely that the majority of adult seroconversions are due to this, as in naturally and experimentally infected kids an antibody response is usually detectable between 3 and 10 weeks following exposure, though delays of up to eight months and possibly longer for seroconversion have been reported (Rimstad et al. 1993).
Additional evidence comes from studies examining the spread of infection in known infected herds. A California study examined the prevalence of seropositive goats in herds attempting to reduce CAE infection by feeding heat- treated colostrum and milk to kids. It was demonstrated that seroconversion still occurred in some of these kids later in life, particularly when CAE-positive adults were maintained in the herd (East et al. 1987). However, when seropositive animals in a herd were physically segregated from seronegative animals that had been raised on heat- treated colostrum and pasteurized milk, rates of seroconversion were threefold lower than when seropositive animals remained commingled with seronegative animals (Rowe et al. 1992).
There are different ways in which horizontal transmission can occur when infected and non-infected dairy goats are commingled. One important route is via milk in association with milking practices. Experimental intramammary transmission of virus has been reported (East et al. 1993; Lerondelle et al. 1995). In one study, direct contact for more than 12 months between uninfected and infected goats was necessary before horizontal transmission could be demonstrated under non-dairy conditions. However, when uninfected does were milked with infected does, 60% became infected in less than 10 months (Adams et al. 1983). Rowe and East (1997) suggest a variety of mechanisms by which horizontal transmission might be effected via milk, including shared milking machines, the impact of backflowing milk against the teat end during machine milking, milk-contaminated hands or towels, aerosolization of milk during milking with subsequent inhalation or contamination of equipment, and spilled or open containers of milk licked or consumed by goats.
In addition to milk, the lungs of infected goats may be a source of aerosolized virus because the lungs are a target organ of CAE infection. Coughing may release virus into the air to be inhaled by nearby animals in confinement housing. Other lung infections may further exacerbate this risk, because bacterial or other viral pneumonias stimulate an inflammatory response that may bring additional CAE virus-laden macrophages to the lungs and increase the frequency of coughing of inflammatory debris. There is good, indirect evidence that aerosol spread of CAE infection is important. In a French study, kids raised on treated colostrum and raw colostrum were maintained in varying degrees of separation and their rates of seroconversion monitored over time. With no separation at all, 34.7% of the kids receiving treated colostrum became seropositive. When separation by a hurdle was imposed, 28.4% serocon- verted. When a wall separated them in the same building, 24.1% seroconverted, and when kept in totally separate buildings, only 16.4% seroconverted (Peretz et al. 1994).
Sexual transmission is a possible but unlikely factor in the spread of CAE infection. There are no reports of infection of does by infected bucks through mating or use of artificial insemination, although in one study does were monitored for seroconversion for 18 months following mating with or artificial insemination from infected bucks (Adams et al. 1983). Epidemiologic studies suggest that the presence of seropositive bucks is a risk factor for the spread of infection in herds, but this is considered to be due to increased direct contact with does during the breeding season rather than through venereal transmission (Kaba et al. 2013). Nevertheless, CAE proviral DNA and viral mRNA have been identified in non-spermatazoan cells of seminal fluid and in various tissues of the genital tract in both experimentally and naturally infected bucks using PCR and in situ hybridization techniques (Travassos et al. 1998; Ali al Ahmad et al. 2008b).
In a similar study, MV virus was found in epididymal tissues in rams coinfected with Brucella ovis and the virus (Preziuso et al. 2003). While the use of only non-infected bucks in breeding programs would be ideal, these observations and findings suggest that CAE virus-infected bucks may be used safely for breeding if some precautions are observed. Infected bucks should not commingle with does, but rather should be hand-mated to does to limit their contact time. They also should be free of any concurrent disease of the genital tract that would possibly increase the presence of virus-infected inflammatory cells in the semen.
There are no reports of CAE transmission associated with embryo transfer. Nevertheless, while embryos with the zona pellucida intact could not be experimentally infected with CAE virus, those with the zona pellucida absent could be infected (Lamara et al. 2002) and supported productive replication of the CAE virus (Ali al Ahmad et al. 2006). In another study, the presence of CAE- infected cells in oviductal washing fluid collected during embryo harvest from infected does was confirmed using a nested PCR for proviral DNA (Fieni et al. 2002). However, a later study demonstrated that embryo transfer can be safely used to produce CAE virus-free neonates from infected CAE virus donors when the embryos are processed according to the guidelines of the International Embryo Transfer Society (Ali al Ahmad et al. 2008a). Nevertheless, for trade purposes, some countries may require that embryos are derived from seronegative flocks.
Iatrogenic transfer of CAE virus is also a consideration in horizontal transmission, though actual occurrences are not well documented. Since the virus is present in peripheral blood mononuclear cells (PMBC), transfer of blood between animals by multiple use of needles for injection and the use of tattooing instruments without disinfection between animals are potential means of iatrogenic infection.
Finally, a number of behavioral traits were observed in CAE virus-infected dairy goat herds that were considered to contribute, at least potentially, to the spread of infection within the herds (Greenwood et al. 1995). These included teat sucking and biting by does, eye licking in hot weather, contact of the nose and mouth with vaginal and anal areas, nasal secretions present on muzzles of goats and in feed troughs, expulsion of milk into the environment when does lie on their udders, leaking of milk from udders before milking, drinking of milk in strip cups in the milking parlor, and drinking of urine and anal intercourse by bucks. While the risk associated with these different behaviors varies markedly, taken together they underscore both the challenges and the vigilance necessary for implementing successful control programs in infected herds.
Pathogenesis
Lentivirus infections are “slow virus” infections and are characterized by a high prevalence of inapparent infection, a prolonged but variable prepatent period, persistence of the virus in the host, multiple organ system involvement, and a chronic disease course with recurrent episodes of acute disease, such as is seen in the arthritic form of CAE (Cheevers and McGuire 1988).
Persistence of the virus in the host is facilitated by the ability of the lentivirus to sequester as provirus in host cells without the necessity of frequent multiplication. The CAE virus uses the monocyte/macrophage cell line as its principal host cell type. The virus remains inactive in monocytes and replication of virus is linked with maturation of monocytes to macrophages after they leave the bone marrow or blood and localize in tissue sites. This has been referred to as a “Trojan horse” strategy, whereby virus gains access to target tissues undetected in monocytes (Peluso et al. 1985). Activation of disease is associated with activation of virus translational activity in infected cells. The mechanisms by which lentiviral replication is first restricted and then subsequently expressed are complex and not yet completely understood, but the subject has been reviewed (Bertoni 2007). An important factor may be availability of viral transcription factors present in mature, tissue-bound macrophages, but not present in circulating monocytes.
Infection of the goat with CAE virus induces both a strong humoral and cell-mediated immune response, but neither is protective. In fact, CAE is an immunopathologic disease in which lesions result from an immune reaction to viral antigens, especially surface glycoproteins. This concept is supported by numerous observations, including the fact that vaccinated goats develop more severe lesions when experimentally challenged than do unvaccinated goats, that persistently infected goats develop acute arthritis when experimentally challenged, that the severity of joint lesions correlates with the presence of virus and antiviral antibody in the joint, and that the lesions occurring in CAE contain mainly lymphocytes and macrophages (Adams et al. 1985; McGuire et al. 1986).
After kids consume colostrum or milk containing CAE virus-infected macrophages, these infected cells are taken up intact from the gut and enter the reticulendothelial system to establish infection in the kid (Narayan and Cork 1985). Subsequently, infected monocytes reach target tissues such as synovium, lung interstitium, choroid plexus, and udder, where the activation of virus replication in conjunction with macrophage maturation induces the lymphoproliferative lesions that characterize CAE. Lungworms may possibly predispose to the pneumonic form of CAE in infected goats by inducing monocyte migration and macrophage proliferation in parasitized lung (Ellis et al. 1988).
Maternal antibody against CAE provides no protection against infection in suckling neonates and usually disappears within 60-85 days after suckling (Adams et al. 1983). Kids infected at birth can show an active antibody response as early as 3-4 weeks and usually by 10 weeks (Ellis et al. 1986). When the antibody response develops, antibodies are present for life, although the serum concentration may wax and wane at different times or may be below the threshold of tests used to detect them.
Clinical Findings
In addition to subclinical infection, there are five known clinical forms of CAE in goats: arthritis, leukoencephalo- myelitis, interstitial pneumonia, mastitis, and progressive weight loss. Arthritis, the most common form, is discussed here. The neurologic form is discussed in detail in Chapter 5 in association with MV. The respiratory form, a chronic progressive interstitial pneumonia of adult goats, is described in Chapter 9. It is sometimes also observed clinically in goats with the arthritic form. The udder form, an indurative viral mastitis with hypogalactia or agalactia noted at parturition in young does, is described further in Chapter 14. Progressive weight loss can occur as the sole clinical manifestation of CAE virus infection, or in conjunction with any of these other clinical forms. It is discussed in more detail in Chapter 15. Enlargement of associated regional lymph nodes occurs with the various clinical forms of CAE virus infection. The arthritic form of CAE occurs in sexually mature goats, usually after the first year, most commonly in the second, but at any time in adulthood. The onset may be insidious or acute and the clinical course varies considerably among individuals. All limb joints and the atlanto-occipital bursa may be affected. The most common site is the carpal joint; the next most commonly involved joints are the tarsal, stifle, and fetlock joints; the atlanto-occipital bursa; and finally the coxofem- oral joint. A single joint or multiple joints may be involved at any time in the course of disease.
Early signs of arthritis may be subtle. Joints may appear and feel normal, but affected goats may have decreased ambulation and feeding activity, a reluctance or difficulty in rising, a stiff gait or abnormal posture after rising, and weight loss. Alternatively, affected goats early in the course of disease may present with an obvious, acute swelling in association with the joint, but with little evidence of pain or restricted activity.
The most obvious swellings are on the anterior aspect of the carpus and are periarticular, associated with fluid accumulating in the carpal bursa. Such swellings may be up to 10 cm in diameter, and are fluctuant, cool, and not painful to the touch (Figure 4.6a). Fluid can be aspirated from these swellings for diagnostic purposes, but drainage only leads to refilling. The condition may remain at this stage of development in some animals for months or even years, or
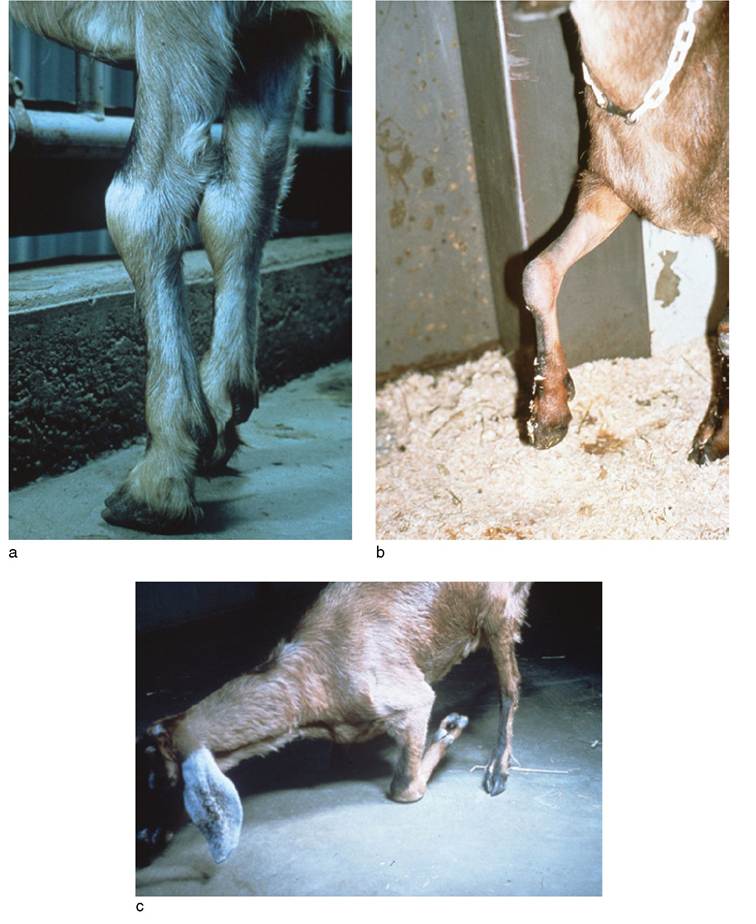
Figure 4.6 Clinical progression of goats with carpal arthritis due to caprine arthritis encephalitis virus (CAEV). (a) Typical carpal swelling in early stage of CAEV- induced arthritis. Reproduced by permission of Dr. C.S.F. Williams. (b) Partial flexure contraction of the forelimb in a goat with progressive CAEV- induced arthritis of the carpus. Source: Courtesy of Dr. David M. Sherman. (c) Permanent kneeling due to ankylosis of the carpi in a goat with advanced CAEV arthritis of the carpi. Source: Reproduced by permission of Dr. C.S.F. Williams.
show periodic cycles of swelling, possible pain, and spontaneous improvement.
For many goats, however, the infection progresses steadily to a debilitating and painful arthritis. This is usually accompanied by gradual weight loss and a rough haircoat. Fevers are not part of the disease process. The fluctuant swelling of the bursa gives way to a firmer swelling associated with inflammation and thickening of the joint capsule and surrounding structures, and the animal becomes increasingly reluctant to use the affected limb. Progressive mineralization of the joint capsule and soft tissues, exostoses of the bones, and collapse of the joint contribute to pain, a restricted range of motion, and finally ankylosis. When the carpus is involved, there is a marked tendency to carry the limb in a partially flexed position (Figure 4.6b). Ultimately, a permanent, severe flexion of the carpus may occur, and when both forelimbs are involved, the animal is forced to ambulate in a permanently kneeling position on the carpi (Figure 4.6c).
Involvement of the hindlimbs usually results in less obvious swellings, though thickening of periarticular structures may be palpable in the hock region. There is progressive reluctance to rise and increasing stiffness of gait. When the hips are involved the animal may develop a sufficiently painful and swaying gait to erroneously suggest neurogenic incoordination.
Clinical Pathology and Necropsy
As CAE virus infection is persistent in the host, virus isolation is not routinely conducted for diagnostic purposes, as confirmation of the presence of antibody infers the presence of virus. In cases where CAE is strongly suspected but seroconversion has not yet occurred, the CAE virus can be identified from live animals by in vitro co-cultivation of concentrated leukocyte preparations derived from blood, milk, or synovial fluid with goat synovial membrane in cell culture. At necropsy, suspect tissues from joint, lung, or udder can be cultivated directly in tissue culture flasks and examined for cytopathic effect (CPE), which is manifested as the development of refractile stellate cells and syncytia formation. When CPE is seen, the presence of virus should be confirmed by immunolabeling or electron microscopy (OIE 2018e).
Most laboratory testing conducted in regard to CAE is for the purposes of surveillance and implementation of control programs and requires the identification of clinically and subclinically infected animals, mainly through the application of serologic tests. The most commonly employed serologic tests are the AGID, the indirect enzyme-linked immunoassay (iELISA) and the competitive ELISA (cELISA). The AGID and ELISA tests are the recommended tests for use in international trade to determine CAE infection status (OIE 2018e). Additional, supplementary tests for detection of antibody are the radioimmunoprecipitation assay (RIPA), western blot (WB), and radioimmunoassay (RIA).
The AGID historically was most commonly used test for routine screening, providing good sensitivity and excellent specificity. Antigen for the prescribed AGID test is derived from whole virus that has been concentrated from culture supernatants and contains both the core antigen p28 and envelope antigen gp135. Detection of antibodies to both antigens improves the sensitivity of the test compared to using one or the other. While there is cross-reactivity of antibodies against the CAE and MV viruses, the sensitivity of the AGID test is improved if the CAE virus is used to detect CAE infection in goats.
Many different configurations of ELISA tests for detection of SRLV infection have been reported, with considerable variation in their performance relative to the AGID and relative to each other. A summary of all these SRLV ELISA test results has been published (de Andres et al. 2005). While early iterations of ELISA did not perform as reliably as the AGID, improvements in antigen development leading to enhanced sensitivity, along with the fact that ELISA can be automated for large-scale sample processing, have resulted in ELISA becoming the preferred test for many laboratories and regulatory agencies. There are two main categories of ELISA assay in use: antibody-detecting iELISAs, which use either whole virus or recombinant proteins/synthetic peptides as antigens, and cELISAs, based on the use of antiviral monoclonal antibodies. For iELISAs overall, assays using whole virus antigen tend to be more sensitive than single recombinant antigen assays. However, if both a core antigen and an envelope antigen are included in the recombinant antigen assays, sensitivities and specificities are equivalent to whole cell antigen iELISAs (de Andres et al. 2005; OIE 2018e).
In the absence of a gold standard reference test to definitively establish CAE infection, RIPA, WB, and RIA are often used as reference tests to establish the comparative sensitivity and specificity of various serologic methods. They can also be used directly in serosurveillance, but are generally considered too cumbersome and costly for high-throughput testing.
The major limitation in the use of serosurveillance for control of CAE is the fact that delayed seroconversion is known to occur in CAE infection, as discussed earlier. With delays in seroconversion of eight months or longer following infection, some infected individuals will be missed in herd-wide testing programs. One potential solution to this problem is the detection of proviral DNA by use of real-time or quantitative PCR techniques. Various PCR protocols have been reported, with most studies using PMBC as the target cell for CAE virus detection. However, since only approximately one in a million leukocytes contains virus, PCR assays may fail to detect virus because the virus load is below the detection limit of the assay, and some investigators have employed co-cultivation of PMBC with fibroblasts to increase virus load. Overall, PCR tests have tended to be less sensitive than many ELISA techniques (de Andres et al. 2005). Nevertheless, there are reports of PCR identifying CAE virus-infected goats prior to seroconversion without co-cultivation (Reddy et al. 1993; Rimstad et al. 1993).
In addition to testing of PBMC, PCR has been applied with equivalent results to semen, milk, and synovial fluid of goats. Currently, it is recommended that serology be combined with PCR for optimal detection of SRLV infection (de Andres et al. 2005). Loop-mediated isothermal amplification (LAMP) also has been reported for the detection of CAE provirus from blood samples (Balbin et al. 2014, 2017).
When seeking to confirm clinical cases of CAE, care must be taken in interpreting positive serology. As there are many causes of arthritis, pneumonia, and mastitis in goats, and because the majority of CAE-infected animals are subclinically infected, positive serology for CAE does not necessarily mean that the swollen joint, respiratory signs, or abnormal udder are due to CAE infection.
In the case of arthritis, arthrocentesis and examination of synovial fluid can support the diagnosis of CAE-induced arthritis. In active cases, the fluid is red to brown in color and has a low viscosity. Cell counts are elevated to the range of 1000-20 000/mm3, with mononuclear cells comprising more than 90% of the cell population. The majority are lymphocytes and the remainder are synovial cells and macrophages (Crawford and Adams 1981). Synovial biopsy can also be useful for histopathologic examination or for virus culture by explantation techniques. Examination of cells in synovial fluid using PCR techniques may also aid in the diagnosis when lesions and clinical signs are consistent with CAE but serology is negative (Reddy et al. 1993).
A wide range of radiographic findings can be observed. In early cases, distension of bursae with fluid and other periarticular soft tissue swellings may be present. In advanced cases, CAE arthritis is characterized by more severe soft tissue changes, including calcification of joint capsule and adjacent tendons, tendon sheaths, ligaments, and bursal contents (Figure 4.7). Early bony changes include mild periosteal reactions and periarticular osteophyte production. Later signs are more severe and include marked osteophyte production, breakdown of subchondral bone, collapse of joint spaces, and ankylosis (Crawford and Adams 1981).
At necropsy, affected goats are usually emaciated. Affected joints and adjacent bursae and tendon sheaths are usually enlarged. Foci of mineralization and/or necrosis as well as fibrous scar tissue may be visible in tendons and other periarticular structures. When the joint is opened, a dramatic synovial proliferation is frequently observed. The synovial membrane appears brown and velvety and is thrown up into fingerlike projections. Cartilage surfaces can be roughened, ulcerated, or eroded depending on the severity of the case. “Rice bodies” are commonly found in the joint space, tendon sheaths, and bursae. Collapse of subchondral bone and ankylosis may also be noted (Crawford and Adams 1981).
Histologically, joint lesions are characterized by synovial hyperplasia with subsynovial mononuclear infiltrates, including lymphocytes, macrophages, and plasma cells. Multinucleated cells are occasionally observed. Fibrin
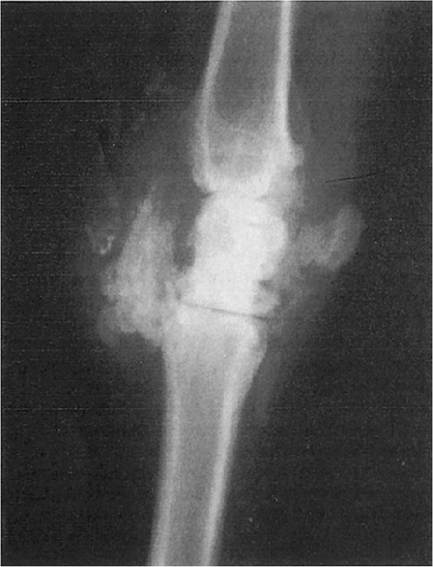
Figure 4.7 Radiographic changes in the carpus of a goat with advanced caprine arthritis encephalitis arthritis. Note mineralization of extensor tendon and other soft tissue structures. Source: Courtesy of Cummings School of Veterinary Medicine at Tufts University.
deposition is common on synovial surfaces. There is necrosis and mineralization of synovial, perisynovial, and tendon collagen. The lesion of mononuclear infiltration seen in the synovium may also be seen in other target tissues such as udder and lung, even when there has been no clinical involvement of these organs. Similar mononuclear infiltrative lesions have also been observed in the uterus of a CAE virus-infected goat (Ali 1987) and the kidney of another (Dawson et al. 1983). Central nervous system lesions are described in Chapter 5, lung lesions in Chapter 9, and udder lesions in Chapter 14.
Diagnosis
A presumptive diagnosis of CAE arthritis is based on a combination of a history of CAE in the herd, positive serology and/or PCR, clinical signs consistent with the disease, synovial fluid abnormalities consistent with the disease, and typical histopathology on synovial biopsy or postmortem examination. All other traumatic, metabolic, and infectious causes of arthritis must be differentiated from CAE arthritis. Concurrent processes can occur, so cultures of synovial fluid for mycoplasma and bacteria are advised for definitive case assessment.
Treatment
There are no known treatments for any of the clinical forms of CAE. Animals with mild cases of the arthritic form of the disease can be made more comfortable by providing regular, correct hoof trimming, by providing easily accessible feed and water, and by the long-term use of oral analgesics such as aspirin, phenylbutazone, or meloxi- cam. Aspirin can be given at an oral dose of 100 mg/kg every 12 hours, phenylbutazone at an oral dose of 10 mg/ kg once a day, or meloxicam at an oral dose of 1 mg/kg once a day. These drugs should be avoided in goats pro - ducing meat or milk for human consumption. Additional non-steroidal anti-inflammatory drugs are discussed in Chapter 17. Physical therapy might delay the progress of CAE arthritis, but there is no published evidence to support this.
In the case of goats with severe, advanced arthritis, which are unable to extend their legs and are forced to walk on their flexed carpi, euthanasia should be considered. Such goats are less frequently encountered nowadays than they were when the disease was first recognized, because of increased awareness on the part of owners about CAE and more aggressive culling of clinical cases. Discovering such goats on a farm should alert the veterinarian to the need for client education concerning CAE.
Control
Attempts to prevent the introduction of CAE into CAE-free countries or zones are based on import regulations that require validation of negative status in individual animals to be imported. The international standard suggested by the World Organization for Animal Health (OIE) is that animals be accompanied by an international veterinary certificate attesting that the animals showed no clinical sign of CAE on the day of shipment, that animals over 1 year of age were subjected to a diagnostic test for CAE with negative results during the 30 days prior to shipment, or that CAE was neither clinically nor serologically diagnosed in the sheep and goats present in the flocks of origin during the past three years, and also that no sheep or goat from a flock of inferior health status was introduced into these flocks during that period. The recommended serologic test is AGID or ELISA (OIE 2018e).
For countries, zones, or states where CAE is already present, testing and certification/accreditation programs may be implemented for control. Few countries have mandatory, nationwide regulatory control programs for CAE. Switzerland is one, having implemented a mandatory CAE control program in the 1980s, which led to successful eradication of the disease. Nevertheless, Switzerland remains vigilant and monitoring continues in order to prevent reintroduction through cross-border livestock movements (Martin et al. 2019).
Other countries, for example New Zealand, France, and Sweden, have instituted voluntary testing and accreditation programs, usually in association with producer groups to encourage the establishment of CAE-negative herds and flocks (MacDiarmid 1985; Davidson 2002; K. de Verdier, Department of Animal Health and Antibiotic Strategies. National Veterinary Institute (SVA), Uppsala, Sweden, personal communication, April 2008; C. Chartier, Director, Agence Franqaise de Securite Sanitaire des Aliments, personal communication, April 2008). Australia has voluntary programs operating on the state and territory level (Animal Health Australia 2009). Such control programs usually involve annual or semiannual, herd-wide serologic testing and removal of positive reactors from the herd. The European Union (EU) does not require compulsory control programs for its member countries, but control/eradication programs can be mandated within EU countries. One reported example involves the Autonomous Province of Bolzano - South Tyrol in Italy (Tavella et al. 2018). In the United States, there is no CAE control program at the national level, though various states, e.g. Minnesota, may have voluntary programs.
At the farm level, efforts to eliminate CAE from herds known to be infected focus on three main areas: attempting to raise CAE-free kids by implementing kid-rearing techniques that reduce the risk of exposure to virus; periodic serologic testing of the herd; and culling or separating diseased and seropositive animals from healthy, seronegative animals. There are no vaccines available for the control of CAE.
Raising Caprine Arthritis Encephalitis-Free Kids
Modification of kid-rearing techniques is probably the most frequently implemented aspect of herd control programs. The goal is to eliminate exposure of kids to virus and is based on the assumption that most new infections in kids develop in an infected herd as a result of ingestion of virus-contaminated milk or colostrum. More stringent programs also remove kids from dams immediately at birth, on the assumption that the potential exists for kids to be exposed to virus when they are licked and cleaned by their infected dams. Washing birth fluids off the kids immediately after birth has also been recommended.
To achieve this level of control, attendants need to be present at birth so kids can be removed from their dams before they are able to stand and suckle. To improve the chance of being present at kidding, timed induction of parturition with prostaglandins can be used, as discussed in Chapter 13. Once kids are removed from the doe, they should be raised in separate facilities from adult animals, especially if infected adults are maintained on the farm, and they should never again be commingled with known infected goats.
Ideally, kids removed from the dam should be fed heat- treated goat colostrum as an appropriate, CAE virus-free source of maternal antibodies to provide passive immunity against neonatal diseases. Pasteurization of colostrum at high temperature for short periods can denature protective immunoglobulins and causes the colostrum to gel, making it impossible to feed. As an alternative, heat treatment of colostrum at 56 °C (132.8 °F) for 60 minutes has been demonstrated experimentally to block transmission of virus and permit absorption of immunoglobulins (Adams et al. 1983).
Automated equipment for performing this task is now available, but may not be affordable for many small herds. Owners of small herds can be advised to heat colostrum to 57 °C (134.6 °F) for 10 minutes and then transfer it to a thermos flask that has been preheated with boiling water. The colostrum is then held in the closed thermos flask for 60 minutes to accomplish virus inactivation. It is essential that the temperature remain above 56 °C (132.8 °F) for the entire period (MacKenzie et al. 1987) and it should be verified by thermometer at the end of the holding period. Some producers are now using multicookers (Instant Pot®, Instant Brands, Downers Grove, IL, USA) to hold the colostrum at the appropriate temperature for one hour. Excess colostrum may be successfully frozen in small aliquots for later use to reduce labor expenditure.
Because heat treatment of colostrum is a laborious task, goat owners actively seek alternatives to this practice. Feeding no colostrum should be viewed as an unacceptable practice, because this results in failure of transfer of passive immunity (FTPI) and puts kids at risk for other dangerous diseases. Feeding unpasteurized colostrum only from seronegative does is not without risk, because falsenegative serologic test results do occur. Administration of unpasteurized cow colostrum results in acceptable levels of circulating serum antibodies in kids, but the persistence and efficacy of such heterologous antibody in kids have not been documented. The practice is reported to be effective in raising orphan lambs. Of course, unpasteurized cow colostrum, while it does not contain CAE virus, may contain other organisms such as Mycobacterium paratuberculosis that are potentially infectious for goats. In addition, feeding of cow colostrum has occasionally been associated with hemolytic anemia in kids (Perrin and Polack 1988).
A number of products are being marketed as colostrum supplements or replacers for calves, lambs, or kids. Such products usually contain lyophilized bovine colostrum or whey immunoglobulin concentrates. Many goat owners have seized upon these products as colostrum substitutes suitable for use in CAE control programs. However, it has been demonstrated that some of these products, when administered to kids as directed by the manufacturer, do not raise serum antibody levels above a level consistent with a diagnosis of FTPI (Sherman et al. 1990). Numerous anecdotal reports suggest a marked increase in the incidence of neonatal septicemia when some colostrum supplements have been used in place of colostrum (Scroggs 1989; Custer 1990).
Newer products on the market may be more reliable, but all such products should be used with caution and kids monitored for an increased occurrence of neonatal septicemia or other disease. Antibody levels in kid serum can be measured by a number of methods, as discussed in the section on FTPI in Chapter 7.
After administration of heat-treated goat colostrum, kids must be raised on pasteurized goat milk, cow milk, or commercial milk replacer through weaning. Goat milk can be pasteurized by heating to 73.9 °C (165 °F) for 15 seconds. The feeding of milk replacers to kids is discussed in Chapter 19.
Periodic Serologic Testing
Submission of serum samples from all goats of breeding age and older for detection of antibody to CAE virus can be useful to establish the prevalence of disease, monitor progress in control programs, serve as a basis for creating infected and non-infected strings of goats in the herd, or identify animals for culling.
There is not universal agreement on the appropriate frequency of testing for herd control programs and the frequency of testing required varies between herds based on initial level of infection, size of herd, capacity to rigorously implement necessary management changes, and other epidemiologic factors. It has been suggested, for example, that kids be tested monthly up to 6 months of age because of variability in the time of seroconversion following perinatal infection. This is not necessary if the kid-rearing interventions discussed above are assiduously practiced, but if management is lax then such testing may be worthwhile to identify perinatally exposed kids and to underscore lapses in control practices. It should be noted that kids fed heat- treated colostrum from infected does will have falsepositive serologic tests for several months at least.
In general, goats should be tested at least annually, and preferably semiannually, because sensitivity of the serologic tests is not 100% and serum antibody levels may fluctuate in individual goats, as in late pregnancy when serum antibody levels may be reduced due to maternal transfer of antibodies from serum to colostrum. Timing a herd-wide test for the beginning of the breeding season is useful, because the results can be used to advantage in making breeding, culling, and management decisions relating to CAE control well before kidding time.
Culling or Separation
Kids raised CAE free using the above protocols may still seroconvert after weaning as a result of horizontal transmission in herds where known infected adults are allowed to remain and commingle in the herd (East et al. 1987). Herd owners attempting to eliminate CAE from their herds should be strongly encouraged to include aggressive culling of seropositive adults as an integral part of the control program. Otherwise, the efforts placed on rearing of CAE- free kids may, in many cases, be futile.
However, in many infected herds, particularly commercial milking herds where the need for milk and replacement does is high, there is frequently resistance to the notion of culling seropositive adults on economic grounds, even though kid-rearing techniques are practiced to reduce the prevalence of CAE. In these cases, it is necessary to maintain two separate groups within the herd - a seropositive group and a seronegative group - that are physically separated to the greatest extent possible. Separate buildings are ideal, but may be impractical. Solid wall separation is the next best option, but if that too is not possible, then the animals must be physically separated by a distance of at least 2 m with a double fence or barrier that allows no direct contact between individuals in the two groups.
Separate feeding, watering, grooming, and other such equipment should be maintained for each group, but if that is not possible, the equipment should be used first on seronegative animals and always washed and disinfected after use on seropositive animals. Seronegative goats should be milked first as a string before seropositive goats, and considerable care must be taken to avoid contamination of surfaces, clothing, and equipment with milk from seropositive does in milking parlors, where it might make contact with seronegative does the next time they are milked.
Additional Considerations
Other practical considerations for effective control programs are to avoid bringing goats to fairs and shows if possible, or at least to avoid commingling goats either at the event or during transport to and from the event. No animals should be introduced into a herd attempting to eliminate CAE unless that animal comes from a known CAE-negative herd, or the individuals test negative and can be quarantined for six months and retested before entering the herd. Ideally, only seronegative bucks should be used for natural service, but if that is not possible, then the buck should be left with the doe only long enough to breed under observation to avoid unnecessary contact (Kaba et al. 2013). Current information suggests that artificial insemination and embryo transfer can be used in flocks seeking to control CAE as long as the semen and embryos are properly processed (Fieni et al. 2012).
Because the virus is present in peripheral blood monocytes in infected animals, the use of separate needles and syringes for each animal during treatment and vaccination is advised, because iatrogenic transmission is at least theoretically possible. Similarly, tattooing of goats may involve blood contamination of the tattooing instrument, so disinfection with phenolic or quaternary ammonium compounds between uses is advised.
If sheep are kept on the same premises with goats and CAE control efforts are undertaken, sheep should be included in the testing program and subjected to similar segregation and management interventions for control. Recent evidence indicates that cross-transmission of SRLV between the two species occurs (Shah et al. 2004a, b) and may adversely affect CAE control programs if sheep are not included in the program design (Minardi da Cruz et al. 2013; Martin et al. 2019).
Foot and Mouth Disease
FMD, also known as aphthous fever, is a highly contagious viral disease of wild and domestic, cloven-hoofed animals. Although it usually produces low mortality, it does produce high morbidity and represents a disease of extreme economic importance to animal industries worldwide through loss of productivity and constraints on trade in livestock and livestock products.
In the goat, FMD most often assumes a subclinical form, though clinical cases can and do occur. The major concern with caprine FMD is that subclinically infected goats may act as a source of infection for cattle in mixed livestock systems and through movement and international trade. Knowledge of the disease in small ruminants has been reviewed (Sharma 1981; Pay 1988; Barnett and Cox 1999; Kitching and Hughes 2002; Mahmoud et al. 2019).
Etiology
FMD is caused by a non-enveloped, icosahedral, singlestranded RNA virus of the genus Aphthovirus, in the family Picornaviridae. There are seven immunologically distinct foot and mouth disease virus (FMDV) serotypes, each with numerous strains. The seven serotypes are identified as A, O, C, Asia 1, South African Territories (SAT) 1, SAT 2, and SAT 3. Serotype C has not been detected since 2004 and may no longer be circulating.
The development of immunity after natural exposure or vaccination is highly type specific, and to a varying degree strain specific. Constant global surveillance of outbreaks is necessary to monitor changes in virus type prevalence so that appropriate type- and strain-specific vaccine formulations can be prepared and maintained. The global challenges of maintaining appropriate and sufficient vaccine banks and reserves for FMD has been reviewed (Paton et al. 2005; Lombard and Fussel 2007).
FMDV is cultivated in vitro on bovine thyroid, bovine kidney, or other suitable cell lines, and produces a CPE but no inclusion bodies. It is quite resistant to freezing and desiccation, but does not survive high temperatures or extremes in pH very well. Direct sunlight kills it, but organic debris protects it from the sun. It dies off rapidly in muscle tissue of infected animals, but can persist in offal, blood, and bone marrow. High relative humidity prolongs the life of aerosolized virus. Sodium hydroxide, acetic acid, sodium carbonate, and sodium hypochlorite are the most effective disinfectants. The commercial disinfectant Virkon S, which contains multiple ingredients, has come to be widely used as a disinfectant in FMD outbreak control.
Epidemiology
FMD is one of the most highly regulated livestock diseases in the world, with extensive international efforts undertaken to control the spread of the virus through restricted movement of both live animals and animal products. At present, FMD remains enzootic in large areas of Africa, Asia, and the Middle East. Areas currently recognized by the OIE as free of FMD without the use of vaccination include North America, Central America, and the Caribbean, as well as Australia, New Zealand, Japan, and many island nations of the Pacific. Most countries in South America are recognized by the OIE as free of FMD, some with and some without the use of vaccination (OIE 2020). Most of Europe is free of FMD without vaccination, but some countries remain subject to sporadic incursions of the disease.
At present, serotype O is geographically the most widely distributed FMD serotype in the world and a serotype O PanAsia lineage FMDV has been responsible for many outbreaks in recent decades. This lineage originated in India in 1990 and spread through the Middle East, Turkey, and eastern Europe. It then moved eastward into the People's Republic of China in 1999 and then to Taiwan, South Korea, Japan, Mongolia, and far-east Russia. The virus then appeared transiently in South Africa in late 2000 and in the United Kingdom in February 2001, where it produced a devastating outbreak of FMD. It spread from there to Ireland, France, and the Netherlands, before being brought under control (Grubman and Baxt 2004). Goats are believed to have been involved in the spread of type O PanAsia, at least with regard to its introduction into Saudi Arabia when, in the 1990s, that country banned the importation of cattle from India due to concerns about rinderpest, but continued to import Indian goats and sheep. The current, general distribution of FMD serotypes is shown in Table 4.3. Pools represent independently circulating and evolving FMDV genotypes. Within the pools, cycles of emergence and spread occur that usually affect multiple countries in the region (FAO 2018).
More than 70 species of wild and domesticated mammals are susceptible to FMD infection. Cattle and swine are seriously affected by FMD infection, with high morbidity and overt clinical disease, but comparatively low mortality rates. In contrast, the disease in sheep and goats is less pronounced and often remains subclinical (O'Brien 1943; Nazlioglu 1972; Sharma 1981; Dutta et al. 1984). This is not always the case. In one Indian outbreak of FMD in goats due to virus type A22, morbidity was 82.5% and mortality 8.2% (Mishra and Ghei 1983).
Human infection very rarely occurs, and the disease is generally not considered to be zoonotic. However, the virus can survive in the upper respiratory tract of people for 24 hours after exposure and be transmitted subsequently to animals. This has implications for effective disease control in the age of international jet travel.
FMD continues to be a threat to livestock worldwide, largely because the highly contagious virus is readily transmitted under a wide variety of conditions that are difficult to detect. Among these are the shedding of virus by infected animals before the onset of clinical signs or during convalescence; the existence of subclinically infected, convalescent, or even vaccinated animals that can act as carriers; and the presence of virus in improperly processed products containing meat, milk, bone, blood, offal, or skin of infected
Table 4.3 General global distribution of foot and mouth disease serotypes.
| Region | Serotypes | |
| 1 | Southeast, Central, and East Asia | A, Asia 1, and O |
| 2 | South Asia | A, Asia 1, and O |
| 3 | West Eurasia and Middle East | A, Asia 1, and O |
| 4 | Eastern Africa | O, A, SAT 1, SAT 2, and SAT 3 |
| 5 | West and Central Africa | O, A, SAT 1, and SAT 2 |
| 6 | Southern Africa | SAT 1, SAT 2, and SAT 3 |
| 7 | South America | O and A |
Source: Based on FAO 2018.
animals. The potential for disease transmission via semen during either natural service or artificial insemination has not been investigated in goats, although it has been demonstrated to occur in cattle.
In enzootic regions, the primary mode of transmission is by close or direct contact between infected and susceptible animals. This can be via aerosolization of virus present in the secretions and excretions of the infected animal or by entry of virus through cuts and abrasions or mucous membranes. Wind or other mechanical vectors, including veterinarians on farm visits who do not observe proper biosafety measures, can carry the infective virus long distances. Contaminated bedding, feed, and equipment can serve as a source of infection. Goats and sheep are less likely than cattle to become infected by long-distance airborne virus because their lower respiratory volume makes it more difficult to take in infective doses of virus via aerosol (Kitching and Hughes 2002). Mechanical carriage of virus on humans and vehicles was considered a frequent factor in disease spread between flocks in the 2001 UK outbreak. In areas where pastoralism and communal grazing are common, as in many parts of Africa, south Asia, and the Middle East, infected small ruminants contaminate river water, ponds, pastures, shrubs, and other aspects of the environment, contributing to the dissemination and persistence of disease (Uppal 2004). In serosurveys in Morocco, transhumant and nomadic herds had a higher prevalence of FMD than stationary herds (Barnett and Cox 1999).
Subclinically infected goats and sheep can play an important role in transmission of the disease to cattle. Small ruminants were deemed responsible for epizootics of FMD in cattle in Tunisia in 1989, Greece in 1994, Southeast Asia in 1999, and Turkey in 2001 (Kitching and Hughes 2002; Uppal 2004). In many parts of the world, sheep and goats are herded or managed together and traded in commerce together, so it is often difficult to ascribe a specific role for goats in the transmission or spread of FMD as distinct from sheep. However, there are also some documented situations in which goats are specifically identified as the source of new infections. A goat clandestinely brought from Turkey was deemed responsible for a type O outbreak in cattle in Bulgaria in 1991 (Kitching 1998). Goats have also been implicated in an outbreak of FMD in Kuwait with type Asia 1, when infected goats with this strain were imported from Bangladesh (Kitching and Hughes 2002). The index case for the FMD outbreak in the Netherlands in 2001 was a dairy goat/veal calf farm and the first clinical cases were seen in goats. It is believed that the infection was introduced with purchased, imported calves, but because the calves on the premises were housed individually and the goats commingled freely, circulation and expression of the virus occurred first in the goats (Bouma et al. 2003).
Virus has been isolated from nasal secretions of healthyappearing goats 48 hours before the development of clinical FMD (Raghavan and Dutt 1974). Intranasal inoculation of goats with viruses of A, O, or C type produced a 100% infection rate, with virus present in the oropharynx for up to 28 days in 87% of goats, and no cases of severe clinical disease. Lesions that did occur were mild enough to be overlooked by casual observation (McVicar and Sutmoller 1968). Animals in which virus can be recovered from the oropharynx beyond 28 days post infection are defined as carriers. The maximum duration of the carrier state in goats is reported to be four months, compared to nine months in sheep and up to three and a half years in cattle (Alexandersen et al. 2003).
Some ruminant animals, including some goats, may become carriers when exposed to FMDV, even if they are immune as a result of previous vaccination or recovery from infection. The percentage of animals that become carriers is variable and the infectivity titer of virus in oropharyngeal samples is usually low. Excretion is also intermittent, and the titer declines over time. Development and persistence of the carrier state may be determined by both the animal species and the strain of virus involved (Alexandersen et al. 2003). In a review of the role of small ruminants in the transmission of FMD, it was concluded that sheep and goats are most likely to be involved with the transmission of FMDV during the early stages of either clinical or subclinical FMD infection rather than when they are carriers, and that the period of greatest risk of transmission is up to seven days after contact with the infection (Barnett and Cox 1999).
Host species susceptibility may also vary with virus type. For example, a devastating outbreak of FMD in pigs in Taiwan in 1997 with type O virus O/Taw/97 did not cause disease in cattle and goats on the same farms as affected pigs (Yang et al. 1999). That virus strain was related to others involved in earlier porcine FMD outbreaks in the Far East. However, in 1999 FMD occurred again in Taiwan, caused by a type O strain more closely related to type O strains circulating in the Middle East and India, and in the 1999 Taiwan outbreak goats and cattle were clinically affected (Grubman and Baxt 2004).
The frequently mild clinical manifestations of FMD in small ruminants can lead to oversight or misdiagnosis and thereby contribute to the further spread of disease. For example, serotype O was introduced into Tunisia in 1989 in infected sheep and goats imported from the Middle East. Clinical signs were mild, and lameness observed in these animals was initially thought to be due to bluetongue. Eventually the disease spread to cattle, at which time it was quickly identified as FMD due to the more obvious clinical manifestations in that species. However, by then the disease had had time to disseminate and spread into Algeria and Morocco (Kitching 1998). In the United Kingdom in 2001, difficulty in accurately recognizing clinical FMD in sheep contributed to the dissemination of the disease through transport of unrecognized cases. However, it also contributed to the emergency destruction of many sheep flocks deemed clinically affected that subsequently turned out to be negative when test results were completed (Kitching and Hughes 2002).
Pathogenesis
Infection is most commonly initiated by inhalation of aerosolized virus or by direct entry of virus into abraded epithelium of the mouth or feet. After inhalation, multiplication occurs initially in the pharynx and, after direct entry, in the epithelium of the mouth or feet. After 3-11 days, a subsequent viremia ensues in susceptible animals. This is the febrile stage of the disease and lasts three to four days. Virus is found in most tissues and shed in all secretions and excretions during viremia. Young kids may die of viral myocarditis at this stage. In surviving goats, virus ultimately localizes in the epithelial cells of the buccal cavity and feet, and sometimes the teats. Virus multiplication in epithelium leads to hydropic degeneration and coalescence of fluid-filled cells to form enlarging vesicles up to 3 cm or more in diameter in cattle. The overlying epithelium becomes devitalized and the mechanical trauma of eating or walking causes sloughing of the tissue, exposing raw ulcers underneath. This leads to the characteristic signs of stomatitis and lameness when overt clinical disease occurs.
Large, fluid-filled vesicles, such as those seen in cattle, are rarely observed in the mouths of goats and sheep. The thinness of the lingual epithelium in small ruminants causes superficial lesions to rupture early, leaving shallow erosions that usually heal within a few days. Lesions in the mouth of large and, to a lesser extent, small ruminants are most often seen on the dental pad and the tongue, but may also be seen on the lips, gums, and cheeks, and sometimes on the hard palate (Alexandersen et al. 2003).
Clinical Findings
Subclinical infection is the most common outcome in goats exposed to FMD virus. When acute cases occur, initial, non-specific signs of FMD may include dullness, inappetence, restlessness, increased heart and respiratory rates, shivering, and fever. Milking goats may become aga- lactic. Pregnant does may abort. Mortality, when it occurs, is most likely in neonatal kids. Mortality up to 55% in young kids has been observed even when older animals show no morbidity (Hedjazi et al. 1972). In such kids, peracute death from myocarditis often occurs with no other signs observed.
When goats do show specific signs of FMD, they are more often affected on the feet than in the mouth. Therefore, lameness is the most suggestive sign of caprine FMD and demands careful examination of the feet, though oral lesions and signs of stomatitis can and do occur. In a severe Indian outbreak of FMD in goats, 12.5% of cases had only oral lesions, 40% only foot lesions, and 47.5% both foot and oral lesions (Mishra and Ghei 1983). Similarly, in another Indian outbreak in a herd of 385 goats, 8.3% of cases had only oral lesions, 63.8% had only foot lesions, and 27.7% of cases had both foot and oral lesions. The overall morbidity rate was 16% and only one 7-day-old kid died, for a mortality rate of 0.26%. All other affected animals recovered within a period of 10-15 days (Kumar et al. 2004).
Early in the course of clinical disease, vesicles may be found along the coronary band, in the interdigital space, and sometimes on the heels. Any or all of the feet may be involved. Over several days vesicles rupture, leaving behind painful ulcers. Lameness may grow more pronounced and the ulcers on the feet may become secondarily infected with bacteria. Animals may be reluctant to stand or move. Though uncommon, in extreme cases the hoof wall may separate at the coronary band and the hoof slough. Severely affected goats may succumb to starvation, dehydration, predation, septicemia, or secondary pneumonia. In uncomplicated FMD, or with supportive care, oral and foot lesions heal gradually over two to three weeks and goats return to normal, although some animals may remain thin for extended periods.
Early vesicle formation in the oral cavity can lead to signs of lip smacking and drooling. Hyperemic foci or small, raised fluid-filled vesicles may occur in the mucosa of the lips, tongue, and palate, but are likely to rupture quickly due to the thin buccal epithelium in small ruminants. Rupture of oral vesicles exposes mucosal ulcers that may lead to mouth pain and a reluctance to eat, and produces lesions that may be easily confused with other, non- vesicular causes of stomatitis in small ruminants. Vesicles on the teats may also occur (Olah et al. 1976), as may vesicles on the prepuce.
Goats suspected of having FMD need to be inspected very carefully for vesicle formation. Vesicles in the mouth may be small and few in number and not lead to overt signs of stomatitis. Likewise, foot lesions may be mild, affect only one or more limbs, and produce minimal secondary signs. If actual fluid-filled vesicles are not observed, later lesions after vesicle rupture may look merely like abrasions due to trauma.
Even when vesicles occur in the mouth or on the feet of goats with FMD, mild cases may still be overlooked. In a study of experimental infection of goats with a mixture of several strains of SAT 1 serotype of FMDV, goats developed vesicles on the mouth and feet, but the animals did not show a measurable reduction in feeding, suggesting that affected goats did not perceive the oral lesions as a disturbance. Similarly, clinical lameness was not apparent and daily activity, as measured by number of steps with a pedometer, did not consistently decrease in the presence of FMD foot lesions (Lazarus et al. 2019; Wolf et al. 2020). Thus, livestock keepers might overlook clinical FMD in goats during casual observation of their herds. This underscores the need for careful physical examination, including inspection of the feet and oral cavity for detection of caprine FMD, a disease for which early detection and early reporting are critical from a regulatory standpoint.
Clinical Pathology and Necropsy
A definitive diagnosis of FMD from field cases requires laboratory confirmation because the disease cannot be differentiated clinically from other vesicular diseases, including vesicular stomatitis, which can occur, albeit rarely, in goats, and because in small ruminants classical clinical signs as described in cattle may be minimal or absent. Due to the highly contagious nature of this disease and the need to initiate regulatory disease control plans in a timely fashion, laboratory diagnosis of any suspected FMD case is a matter of urgency. When FMD is suspected, regulatory authorities should be notified immediately and sample collection and shipment should be done by trained, authorized personnel according to strict standards of biosafety, with tests performed in a laboratory that meets containment requirements for Biosafety Level Group 4 pathogens.
The best samples to submit for laboratory diagnosis are epithelium from unruptured or freshly ruptured vesicles or vesicular fluid. If such samples are not available, which may commonly be the case in affected goats, blood and/or esophageal-pharyngeal fluid samples taken by probang cup can yield virus. Myocardium or blood can be submitted from fatal cases, but vesicles are preferred when available.
As prescribed by the OIE, diagnosis of FMD is by virus isolation or the demonstration of FMD viral antigens or nucleic acid sequences in tissue or fluid samples. Detection of virus-specific antibody can also be used for diagnosis, and antibodies to viral non-structural proteins (NSPs) can be used to differentiate infection from vaccination. Detailed descriptions and reference sources for laboratory tests used in the diagnosis of FMD are found in the OIE Manual of Diagnostic Tests and Vaccines for Terrestrial Animals, which is available in print and on the internet (OIE 2018a), and are outlined briefly here.
Virus isolation is best performed on cell culture systems using primary bovine (calf) thyroid cells or primary pig, calf, or lamb kidney cells. Other cell lines, such as BHK-21 (baby hamster kidney) and IB-RS-2 cells, may also be used, but are generally less sensitive when there are low amounts of infectivity. CPE occurs in infected cell cultures in 48 hours. If none is seen, cell cultures are frozen and thawed and a new cell culture inoculated to check again for CPE.
For detection of viral antigens, an indirect sandwich ELISA technique has largely replaced complement fixation (CF) as the test of choice in recent years. The ELISA is more specific and sensitive than CF and not affected by pro- or anti-complement factors. This ELISA confirms the presence of FMD virus and also identifies the virus serotype. Lateral flow devices (LFDs) are also now available that can be used to detect FMD viral antigens.
Nucleic acid recognition tests, such as the reverse transcriptase-polymerase chain reaction (RT-PCR), are being used increasingly for virus detection. Agarose gel or real-time RT-PCR can be used to amplify genome fragments of FMD virus directly from epithelium, milk, serum, and oropharyngeal samples or after cell culture of the samples, once a CPE has been observed.
Characterization of FMD strains is very important for tracking the spatial movement of FMD infection, identifying host species adaptations for particular strains, and developing suitable and effective vaccines for specific outbreaks. The molecular epidemiology of FMD is based on the comparison of genetic differences between isolates, using nucleotide sequencing. Dendrograms showing the genomic relationship between vaccine and field strains for all seven serotypes based on sequences derived from the 1D gene, which encodes for the VP1 viral protein, have been produced.
Serologic tests for FMD are conducted for four main reasons: to certify individual animals prior to import or export; to confirm suspected cases of FMD where detection of virus has not been possible; to substantiate the absence of infection; and to demonstrate the efficacy of vaccination. To confirm mild cases or where epithelial tissue cannot be collected, as may often occur in small ruminants, the demonstration of specific antibodies to structural proteins (SPs) of FMD virus in animals known to be unvaccinated is sufficient for a positive diagnosis. The SP tests are serotype specific and detect antibodies elicited by both vaccination and infection. The virus neutralization test, the solid-phase cELISA, and the liquid-phase blocking ELISA for detection of antibodies to SPs are the tests prescribed by the OIE for use in international trade of livestock.
In addition to SPs, FMD virus also expresses eight NSPs during viral replication. There is no viral replication in the host when killed virus vaccines are used for FMD control. Thus, tests for antibodies to some NSPs of FMD virus are useful in providing evidence of previous or current viral replication in the host, independent of vaccination status. In contrast to SPs, NSPs are highly conserved and therefore are not serotype specific, so the detection of these antibodies is not serotype restricted. Unfortunately, some NSPs are produced during growth of vaccine virus in tissue culture, and some traces of these NSPs may contaminate the final killed vaccine product and elicit a host immune response. The NSP 3D is most commonly associated with this contamination and therefore is not a good candidate antigen for NSP antibody tests.
Currently the non-structural polyproteins 3AB and 3ABC are the preferred antigens. Tests prescribed by the OIE for detection of antibodies to NSPs include iELISA and the enzyme-linked immunoelectro-transfer blot assay. The use of diagnostic techniques based on detection of antibodies to NSPs to distinguish FMD virus infection from vaccination has been reviewed (Clavijo et al. 2004).
A full necropsy should be conducted on suspected FMD cases that have died. At the time of necropsy, careful inspection of the feet, oral cavity, pharynx, and teats is required to determine the presence of vesicles or ulcerations consistent with a diagnosis of FMD. In addition, the “tiger heart” lesion may be observed, especially in kids dying peracutely. This name refers to the striped appearance of the myocardium caused by the presence of necrotic, small gray foci and irregular streaks in the heart muscle. These same lesions may occasionally occur in skeletal muscle.
Microscopically, vesicular epithelial lesions include hydropic degeneration of cells in the stratum spinosum, disintegration of cells, and accumulation of fluid. Where vesicles have ruptured, ulcers will be noted and may be associated with suppurative inflammation if secondary bacterial infection has occurred. The heart lesion is one of focal coagulative necrosis accompanied by lymphocytic and sometimes neutrophilic infiltrates.
Diagnosis
Definitive diagnosis is based on isolation of FMDV from suspected cases of FMD detection of viral antigen by ELISA or by detection of viral nucleic acids using molecular techniques such as RT-PCR. Detection of antibodies to NSPs of FMDV is also diagnostic in non-vaccinated animals.
Because vesicular diseases of goats are uncommon, any vesicular lesion of the mouth, muzzle, oral cavity, feet, or teats should be considered as suspect for FMD, even in FMD-free countries, and reported to the appropriate authorities immediately. Because vesicles are fragile and transient in goats, advanced cases may present with ulcers, or crusts over lesions that make the diagnosis less obvious. Therefore, FMD must be considered whenever there are signs of lameness and/or stomatitis, especially when the two signs occur together.
Diseases with the potential of producing both signs together in goats include bluetongue, vesicular stomatitis, pemphigus, and contagious ecthyma. Bluetongue and vesicular stomatitis rarely produce clinical disease in goats, though in the outbreak of bluetongue in Europe in 2006, goats in the Netherlands showed oral lesions and udder lesions (Dercksen et al. 2007). Pemphigus is unlikely to affect more than one goat in a herd. Lesions of contagious ecthyma are found primarily on the mouth, but can be found in other locations as well, including the teats and legs, although overt lameness is unlikely. Ionophore toxicity is an important differential for the myocarditis lesion of FMD that causes sudden death in kids, as discussed in Chapter 8, as is NMD, as discussed later in this chapter.
Treatment
There is no specific treatment for FMD. However, supportive care can help facilitate recovery of affected animals. Such efforts include confinement; provision of easily masticated, high-quality feed and water; and the administration of antibiotics or the application of topical antiseptics to prevent or control secondary bacterial infections of ulcerative lesions. Foot baths using 5% potassium permanganate solution have also been recommended (Kumar et al. 2004). Depending on the regulatory protocols in effect, it may be required to destroy all animals involved in an epizootic, so consideration of treatment may become moot.
Control
Considerable international effort is made to restrict the global spread of FMD and a global FMD control strategy has been developed by the OIE and the Food and Agriculture Organization of the United Nations (FAO) (OIE/FAO 2012). In addition, the OIE, which serves as the standard-setting body of the World Trade Organization with regard to trade in livestock and livestock products, develops and publishes standards aimed at mitigating the risk of FMD in such trade. These standards are available in print or on the internet in the Terrestrial Animal Health Code (OIE 2021). Countries that are FMD free, such as the United States, do not accept imports of livestock from FMD-enzootic countries. However, the importation of certain animal products may be permitted, but only from approved, inspected abattoirs and/or processing plants.
When outbreaks do occur in FMD-free countries, they are eradicated by stamping out procedures and zoo-sanitary measures, with or without the addition of ring vaccination of livestock around the outbreak locale. Countries that are FMD free but that border countries with enzootic infections may practice regular, annual “frontier” vaccination of livestock along the pertinent national boundaries to create buffer zones against incursion of infection.
In enzootic countries, FMD is controlled by restriction of livestock movement, isolation or slaughter of exposed and infected animals, ring vaccination around outbreak locales, or maintenance of a systematic national vaccination program. Effective vaccination depends on ongoing surveillance to ensure that all important, locally occurring strains of FMD virus are included in multivalent vaccines (Blaha 1989). Based on geography, livestock distribution, and other factors, some countries may maintain recognized FMD-free zones within their borders even when the whole country is not considered disease free. The same principles of movement control, surveillance, and vaccination apply for the maintenance of FMD-free zones, and export of livestock and livestock products are permitted from these FMD-free zones when international surveillance and testing standards are met.
Several disease control recommendations were made by the Permanent Commission on Foot and Mouth Disease of the OIE in 1972 specifically concerning goats and sheep (Anonymous 1972). Small ruminants brought to seasonal pasture grounds should be vaccinated against FMD at least once a year. If prophylactic vaccination campaigns are carried out in cattle, then goats and sheep should be vaccinated as well. When it is necessary to undertake ring vaccination of cattle after an outbreak, goats and sheep should be included in the vaccination scheme. Finally, veterinary supervision of goats and sheep should be intensified at markets and during movement to and from pasture grounds, because transhumance, or seasonal movement of livestock to different feeding grounds, has been associated with increased spread of FMD.
Despite these recommendations, inclusion of small ruminants in vaccination efforts is not widely practiced, for several reasons. First, it is not proven that prophylactic vaccination of small ruminants is essential to eradicate the disease in cattle (Barnett and Cox 1999). In addition, there is some evidence that FMD in sheep and goats may be selflimiting (Anderson et al. 1976), although this depends on the strain of virus and the susceptibility of the small ruminants involved, e.g., indigenous breeds versus exotic imports.
Perhaps the most significant resistance to mass vaccination of small ruminants is related to the cost, particularly in developing countries with large populations of sheep and goats and limited veterinary service budgets for disease control. As a result, small ruminants tend to be vaccinated, if at all, only when they are in association with large numbers of cattle or swine. When sheep and goats are included in regular vaccination campaigns in countries where FMD is endemic, they are rarely vaccinated more than once a year (Kitching and Hughes 2002). In Israel, all susceptible species - i.e., cattle, sheep, and goats - are vaccinated regularly every year. In the event of an outbreak, all the animals at the outbreak site and in the surrounding area are immediately revaccinated (Leforban 2002).
Killed vaccines are currently in widest use and are usually trivalent, containing strains A, O, and C. However, many FMD vaccines are produced locally to reflect prevalent serotypes and strains and therefore vary in composition, so blanket recommendations for vaccination cannot be made. Based on local production techniques, they may also vary in immunogenicity, purity, and efficacy. Standards for FMD vaccine production are provided by the OIE (OIE 2018a) and development and performance of killed FMD vaccines has been reviewed (Barteling 2002).
Killed vaccines can be effective in goats when given at one-third the dose volume applied to cattle, with immunity usually lasting from five to six months (Sharma 1981). When cattle, sheep, and goats are vaccinated with inactivated FMD vaccines, sheep, and especially goats, respond with lower mean antibody titers than do cattle (Polydorou et al. 1980; Garland et al. 1981). Optimal responses are obtained when two primary vaccinations are given about two weeks apart, followed by regular boosters at six-month intervals.
The choice of adjuvant used in vaccine may have an impact on goat response to vaccination. In a study comparing oil-adjuvanted and aluminum hydroxide-adjuvanted FMD vaccine, the oil adjuvant elicited superior immune response, the development of the response was quicker, and the persistence of antibody at higher concentrations was greater (Patil et al. 2002). Vaccinating goats for FMD at the same time other vaccines are given does not adversely affect the host response (Gogoi et al. 2004).
Public concerns of animal welfare, sparked by images of the mass destruction of livestock in the United Kingdom during the FMD outbreak of 2001, have stimulated considerable discussion among scientists and regulatory policy makers on the use of vaccination to control FMD as a way of reducing the slaughter of animals. The topic is complex and beyond the scope of this chapter. More extensive discussions of the role of vaccination in FMD control are available elsewhere (LeForban 2002; Pluimers et al. 2002; Constable et al. 2017).
There is considerable ongoing research to develop new and more effective FMD vaccines. Objectives include identification of antigens that might confer immunity across multiple serotypes; vaccine formulations that may stimulate more rapid and stronger protective responses; and
so-called DIVA vaccines, which contain antigenic markers that can be used in serologic testing to distinguish between infected and vaccinated animals. Recent developments in FMD vaccine technology have been reviewed (Barnett and Carabin 2002; Grubman and Baxt 2004; Grubman 2005; Cao et al. 2016). There is also interest in the use of antiviral compounds such as type 1 interferon, which, if administered to animals at risk in an outbreak, can block the establishment of new infections (Grubman 2005).
Legislation enacted in the United States in 2018 established the National Animal Vaccine and Veterinary Countermeasures Bank to enhance preparedness in the event of an outbreak of FMD, which has not occurred in the United States since 1929 (USDA 2020). The vaccine bank will maintain a strategic reserve of frozen vaccine antigen concentrates that can be quickly formulated into a vaccine that reflects the FMDV serotypes and strains involved in a given outbreak. These FMD vaccines will include DIVA technology to be able to distinguish infected from vaccinated animals, which is critical for a country to reestablish FMD-free status if vaccination has been used to successfully control an outbreak.
Akabane Disease and Related Orhthobunyaviruses
Akabane virus, an arthropod-borne, teratogenic orthobun- yavirus, causes congenital arthrogryposis and hydranen- cephaly in kids, lambs, and calves in tropical and temperate regions of Asia, Africa, the Middle East, and Australia. Two other arthropod-borne orhthobunyaviruses are also teratogenic in ruminants, but have different geographic distributions. Cache Valley virus (CVV), first isolated from mosquitoes in Cache Valley, Utah, in 1956, occurs in North, Central, and South America, but clinical disease of small ruminants has only been reported in North America (Waddell et al. 2019). Clinical cases of CVV infection in goats have recently been reported from Iowa (Harvey et al. 2019) and California (UC Davis 2021). Schmallenberg virus was identified in Germany in 2011 and occurs throughout Europe, causing abortions and congenital malformations in calves, lambs, and kids (Collins et al. 2019). CVV and Schmallenberg virus are discussed in more detail in Chapter 13 in relation to abortions.
Etiology
Akabane virus is a helical, enveloped, negative-sense, single-stranded RNA bunyavirus in the Simbu serogroup of the genus Orthobunyavirus in the family Peribunyaviridae, of the order Bunyavirales. The virus can be cultivated in Vero, baby hamster kidney, or hamster lung cell lines. It is an insect-transmitted arbovirus. The principal vector is the biting midge Culicoides brevitarsis, but some other Culicoides spp. and some species of Aedes and Culex mosquitoes are also known to carry the virus. Other arthropod- borne orhthobunyaviruses in the Simbu serogroup, the Aino, Peaton, Sabo, and Shamonda viruses, can also infect ruminants and cause congenital defects, but the occurrence of clinical disease in goats with any of these viruses is not well documented. Antibodies to Tinaroo and Douglas viruses, non-pathogenic members of the Simbu serogroup, have also been reported in goats (Cybinski 1984). An overview of diseases caused by Simbu serogroup orhthobunyaviruses in animals is available elsewhere (Kirkland and St George 2018).
Epidemiology
The host range of Akabane virus includes goats, sheep, cattle, camels, horses, donkeys, and zebras, as well as other wild ruminants. Clinical disease has been observed only in goats, sheep, and cattle. Natural infection of swine has been recorded in Asia, but pigs do not show signs of disease. Congenital arthrogryposis and hydranencephaly of domestic ruminants caused by the Akabane virus have been known in Australia and Japan for decades. Epizootics involving goats, cattle, and sheep have been reported from Japan (Inaba 1979) and Israel (Shimshony 1980). The virus infection is enzootic in Southeast Asia, extending northward to Japan and Korea, southward to northern and eastern Australia, and westward to the Middle East and eastern Africa. In Kenya, 50-90% of cattle sampled had serologic evidence of infection, while prevalence in goats and sheep was only 13-33% (Davies and Jessett 1985). The infection is unknown in the Western Hemisphere.
The geographic pattern of infection reflects closely the habitat of the arthropod vectors, which are Culicoides midges. In Australia, C. brevitarsis is the only certain vector. Akabane virus has been isolated from Culicoides wadai in Australia, but its role as a vector is less well defined. In Japan, Culicoides oxystoma may be the principal vector (Kurogi et al. 1987). New Zealand is free of infection, as Culicoides vectors are not present in the country (Peacock et al. 2018). The virus also has been isolated from the mosquitoes Aedes vexans and Culex tritaeniorhynchus, but mosquitoes are not considered to be true vectors of the virus. In the Middle East, the principal vector is Culicoides imicola (Brenner 2004).
There is a distinct seasonal pattern of virus transmission in countries with a temperate climate. Transmission coincides with warm, moist summer and autumn months, when vector numbers are present in high numbers, and transmission ceases when vector activity subsides with the onset of cold temperatures or frost. In tropical and subtropical regions, a tendency toward seasonal transmission may also be observed, with the highest infection rates in the summer months and transmission rates declining during periods of lower rainfall (Kirkland 2015).
In non-pregnant animals, seroconversion occurs but no disease is observed (Sellers and Herniman 1981). If animals are pregnant, then abortions, stillbirths, or congenital defects can occur. Congenital defects are most likely when dams are infected in early gestation. Malformations in goat kids are common when infection occurs between days 30 and 50 of gestation and kids are normal when infection occurs during the last 100 days of gestation (Kurogi et al. 1977; Shimshony 1980). In temperate zones, goats are seasonal breeders, with conception often occurring in late summer or early fall. As the teratogenic effects observed with Akabane virus infection result from fetal infection mainly in the second month of gestation, this puts goats at potentially high risk for clinically affected offspring.
In endemic areas, annual exposure to the virus through active vectors can produce some level of herd immunity, so that breeding-age females may be resistant to infection at the time of breeding and disease is infrequently noted. However, epizootics may be observed in areas adjacent to endemic areas where the vectors are not regularly present, but may appear when weather conditions permit large populations of Culicoides to develop and expand or be carried by wind beyond their normal distribution limits and nai've populations of livestock are then exposed (Al- Busaidy et al. 1988). New Zealand carries out regular surveillance to detect possible wind-borne incursions of C. brevitarsis from Australia.
Neonatal losses can be severe. In the Israeli epizootic of 1969-1970, up to 50% of the kids born in some herds were affected. Goats appeared more susceptible than sheep, and male, twin kids were more often affected than female, single kids (Shimshony 1980).
Pathogenesis
Infection with Akabane virus does not result in clinical disease unless animals are pregnant. Virus is introduced to susceptible hosts through insect bites. A viremia usually of a two- to four-day duration ensues; in pregnant dams, the virus enters the placenta and fetus. The primary site of viral damage in the fetus is the developing neural tube. In calves, with a bovine gestation of nine months, the outcome of fetal infection depends on the stage of fetal development, with hydranencephaly alone occurring with early infection, hydranencephaly and arthrogryposis occurring in later infection, and arthrogryposis occurring alone in still later infection. Goats have a shorter gestation and, as mentioned above, fetal malformations develop when infection occurs between days 30 and 50 of gestation, and both hydranencephaly and arthrogryposis may be seen together in affected kids.
In hydranencephaly, occlusion of the ependymal system leads to increased cerebrospinal fluid pressure and dilatation of the ventricles, resulting in progressive degeneration and thinning of the cerebral cortex. The cerebellum and brain stem are less frequently affected. Loss of mentation, blindness, and incoordination result from this process. In arthrogryposis, there is a marked diminution or disappearance of ventral horn cells in the spinal cord that leads to a neuromyodysplasia. The failure of normal muscle development in the limbs and along the vertebral column results in muscle dystrophy, tendon contracture, and permanently flexed or extended joints. The joints themselves are normal. Infected fetuses produce antibody to the Akabane virus, and serum samples obtained before the neonate's ingestion of colostrum can be used to diagnose infection.
Clinical Findings
Infection of non-pregnant goats is subclinical. Pregnant goats may remain healthy but abort or deliver stillborn kids, and complaints of abortion may be the first indicator of possible Akabane virus infection. If abortions have not been reported, veterinarians may first encounter Akabane disease when assisting a goat in labor, because dystocias are common when arthrogryposis is present. During epizootics, cattle and sheep in the practice area may also be delivering abnormal calves and lambs. Kids with arthrogryposis are usually born alive and at term. They are unable to rise and affected joints are rigidly flexed or extended. There is severe muscle wasting; there may be scoliosis or wry neck. With hydranencephaly, the kids may be dummies. They may be able to stand and walk if arthrogryposis is not concurrent, but the gait may be uncoordinated. These kids are usually blind and unaware of their surroundings. If a nipple or teat is placed in the mouth, they may suck.
Clinical Pathology and Necropsy
Virus isolation is usually not performed for diagnosis of clinical cases, with serology and histopathology being the preferred methods for confirming infection. Umbilical and heart blood samples or thoracic fluid from aborted fetuses, or precolostral blood samples from live kids, should be tested for antibody to Akabane virus. Virus neutralization titers of 1 : 2 or more are considered evidence of fetal infection and the mean titer of infected goats tested in Israel was 1 : 18 (Kalmar et al. 1975). Titers measured using the hemagglutination inhibition test correlate closely with serum neutralization test results (Furuya et al. 1980). ELISA tests are also in use (OIE 2018b). If aborted fetuses or precolostral serum in kids are not available, titers in does or surviving kids will document previous viral infection in the herd. Because the infection will have occurred months earlier, a positive titer will support but not confirm the virus as the cause of abortions or malformation, and an additional rise in titer is unlikely to occur.
Gross necropsy findings of hydranencephaly are a marked thinning or total disintegration of the cerebral cortex. Occasionally, cerebellar hypoplasia is also observed. In arthrogryposis, the joints are inflexible due to tendon contracture and the muscle bundles surrounding the long bones may seem absent or significantly reduced in volume. The muscles may appear fibrotic and grayish white. If tendons around the joints are cut, the joints move freely and there are no lesions on articular surfaces. Scoliosis, kyphosis, and torticollis may be observed.
Microscopically, there is muscular dystrophy with loss of striation and disappearance of muscle fibers with replacement by fat and connective tissue. Lesions in the spinal cord include a dilated central canal and ventral fissure, cavitation of the ventral gray horns with neuronal degeneration, and edema. In the cerebrum, there is hydrocephalus or more extreme hydranencephaly, subependymal gliosis, and generalized edema. Edema may also be prominent in the cerebellum, and Purkinje cell degeneration is common (Nobel et al. 1971).
Virus may be present in tissues of affected kids or aborted fetuses, especially in the central nervous system and muscles (Kurogi et al. 1977). However, virus isolation is not usually successful because neutralizing antibody is also present in the kid. Alternately, viral antigen may be detected in formalin-fixed fetal tissue by peroxidase staining.
Diagnosis
Akabane disease is diagnosed on the basis of arthrogryposis and/or hydranencephaly in newborn kids and the identification of a positive antibody titer in those kids or aborted fetuses. The possibility remains that other orhthobunyaviruses that produce similar congenital disease in cattle and sheep may be reported to do so as well in goats. In fact, CVV infection recently has been confirmed in deformed goat kids in Iowa (Harvey et al. 2019), California (UC Davis 2021), and another, unpublished outbreak in New York in the winter of 2015, as discussed further in Chapter 13. Currently, the differential diagnosis for congenital arthrogryposis-hydranencephaly in goats is limited to beta mannosidosis, the inherited lysosomal storage disease that can produce arthrogryposis but not hydranen- cephaly and occurs only in Nubian or Nubian-crossbred goats. Bluetongue is an important consideration in sheep, but goats, while frequently seropositive for bluetongue, rarely develop clinical disease. If they do, adult animals are also likely to show signs. As bluetongue is also transmitted by Culicoides midges, Akabane disease and bluetongue may both occur where their vectors are present.
Treatment and Control
There is no treatment for Akabane or CVV infection. Affected kids do not survive long without a good deal of nursing care. There is no practical way to control the insect vectors, although breeding could be delayed until after frosts have killed them in temperate regions. However, effective killed Akabane virus vaccines have been developed and are used in Australia and Japan, where epizootics have previously occurred. Vaccination should be carried out in advance of the breeding season. The killed vaccines are suitable to use for emergency vaccination of pregnant animals (Kirkland 2015).