The White-Nose Syndrome
13.3.1 ClinicalSigns
Due to its psychrophilic temperature requirements, P. destructans can only infect hibernating bats, while in their active life in summer, bats can discard the infection.
In winter during the hibernation period, bats observed at sites affected by WNS display a number of clinical signs either concurrent or singly, but these concern mostly aberrant hibernation behaviour:(a) White powdery fungus, especially on the bats' nose but also on the wings, ears or tail.
(b) Bats flying outside during the day in temperatures at or below zero.
(c) Shifts of large numbers of bats to locations near the hibernacula entrances or unusually cold areas.
(d) Dead or dying bats on the ground or on buildings, trees or other structures. On occasion, large numbers of fatalities occur either inside the hibernacula, in the immediate vicinity of the entrance.
(e) Bats show a general unresponsiveness to human disturbance.
(f) Bats arouse with markedly increased frequency from torpor (survival strategy where metabolism is reduced to a minimum) (Reeder et al. 2012; Warnecke et al. 2012).
(g) WNS-affected bats groom themselves in higher rates, with particular focus on their wing membranes (Brownlee-Bouboulis and Reeder 2013).
13.3.2 HistopathologicalChanges
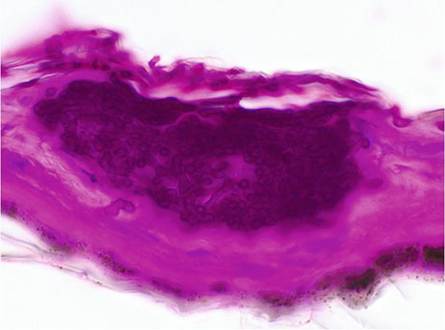
The infection of hibernating bats with P. destructans is restricted to the glabrous skin of muzzle, ears and wing membranes, but in contrast to most dermatophytes, P. destructans invades the epidermis deep into the underlying connective tissue (Meteyer et al. 2009). Hallmarks of the disease are so-called cupping erosions in the epidermis containing dense pockets of fungal hyphae (Fig. 13.5) and/or ulcerations caused by fungal hyphae penetrating into the dermis, sometimes spanning the whole thickness of the wing membrane (Meteyer et al. 2009). At the muzzle fungal hyphae usually fill hair follicles and invade sebaceous and apocrine glands while also reaching into the adjacent connective tissue.
Hibernating North American bats have no inflammatory response regardless of the severity of the lesions. This is attributed to the hibernation status where all body functions are reduced to a minimum including the immune system (Bouma et al. 2010). However, North American bats with severe wing damage, which were caught shortly after hibernation, as well as European bats in late hibernation revealed influx of neutrophilic granulocytes and macrophages as well as intradermal abscesses (Meteyer et al. 2009; Wibbelt et al. 2013; Zukal et al. 2014). Whether the differences in inflammatoryFig. 13.5 Lightmicroscopy image of epidermal “cupping erosion” caused by hyphae of Pseudogymnoascus destructans (PAS stain)
response between hibernating North American and European bats are merely due to the time of sampling with regard to hibernation length or a true species difference is currently unclear.
13.3.3 Pathogenesis
Despite the successful proof of P. destructans as the causative agent for WNS lesions in infection experiments (Lorch et al. 2011; Warnecke et al. 2013), the exact pathogenesis of the deadly outcome of this fungal infection on North American bats still remains unclear.
First investigations of deceased bats reported emaciated body conditions in little brown bats (Myotis Iucifugus) (Meteyer et al. 2009; Courtin et al. 2010). An extensive study on free-ranging little brown bats used temperature-sensitive data logger fixed to the back of individual bats to investigate the frequency of arousal from torpor bouts, as during arousal bats will resume euthermic body temperature for a short period of time. Evidently, bats affected by P. destructans or bats that died due to the infection aroused significantly more often than unaffected bats and had shortened torpor bouts (Reeder et al. 2012). The same results were drawn from experimental infections, where two groups of little brown bats infected with a North American and a European fungal strain, respectively, presented a markedly increased arousal pattern (Warnecke et al.
2012). As arousals involve a high energy demand, it is thought that WNS-affected bats most likely burn their fuel storage prematurely before the end of winter and subsequently die of starvation. Additionally many WNS-positive bats are severely dehydrated. Current hypotheses assume that either due to increased arousals more time is spent in an euthermic, waterconsuming state or that severe epidermal damage by P. destructans disrupts the wing membrane integrity. As the wing membranes play an important part in homeostasis and water balance, this could result in increased evaporative water loss (Cryan et al. 2010). The small body size of little brown bats (5-14 g body weight) is one of the many obstacles to answer these questions, for example, through the analysis of blood parameters. A number of investigations attempted interpretation of blood values from P. destructans-infected free-ranging as well as from captive little brown bats (Warnecke et al. 2013; Cryan et al. 2013; Verant et al. 2014), but results partly contradicted each other and did not satisfactorily explain the mechanisms behind the cause of death in WNS-affected bats.European bats sometimes display large areas of wing membrane covered with numerous fungal colonies (Puechmaille et al. 2011; Wibbelt et al. 2013; Zukal et al. 2016) and still seem in good body condition. Bats from China had vastly reduced fungal loads compared to North American species and were similarly thriving as their European conspecifics. These circumstances could indicate a possible host resistance of these bat species being exposed to P. destructans for very long time (Hoyt et al. 2016). A thorough study of the transcriptome of WNS-affected and WNS-unaffected North American bats found numerous upregulated genes responsible for inflammatory responses in affected bats, but nevertheless local inflammatory response recruitment of leukocytes to the infection sites is missing (Field et al. 2015). Additionally, the transcriptome of the fungus was retrieved, detecting several putative virulence factors indicating host-pathogen interactions.
Still the retrieved data cannot fully explain the complex mechanisms behind the pathogenesis.An interesting approach is a new modelling study, which takes the environmental requirements of P. destructans, i.e. growth temperature and relative humidity, as well as hibernation requirements of different bat species into account. The results are surprisingly similar to observations from field studies and experimental infections and suggest that environmental conditions and basic host traits alone may explain much of the variability in disease outcomes among species of bats infected by P. destructans in North America and Europe. Moreover, the key features of fungal ecology are inextricably linked to bat mortality from WNS (Hayman et al. 2016).
13.3.4 SamplingandDiagnosis
13.3.4.1 Time of Sampling
P. destructans is a very slow-growing fungus, and colonies on the skin of hibernating bats will most likely be visible only when several weeks of the hibernation period have already passed. Sampling efforts should be attempted about 3 months after bats entered hibernation. The later towards the end of hibernation, the higher the chance that aerial mycelia have accumulated in visible patches. However, if bats start to arise from hibernation, they can quickly groom off all visible traces (Puechmaille et al. 2011).
13.3.4.2 Touch Imprints
Loops of adhesive tape can be used for quick analysis of suspicious fungal infection. Lowering the sticky side of a tape loop towards the surface of a fungal patch on

Fig. 13.6 Light microscopy image of unstained touch imprint from Pseudogymnoascus destructans (left); remaining adhesive tape of touch imprint after being dissected for culture or PCR use (right)
hibernating bats allows enough material to adhere (Meteyer et al. 2009; Wibbelt et al. 2010). Immediate transfer onto a glass slide and examination by light microscopy with narrowed condenser aperture and 100 times magnification make P. destructans' characteristic conidia easily visible (Fig. 13.6). After thorough cleansing of the tape's outer surface with 96% ethanol, cut-out areas laden with conidia can be used for isolation of pure cultures and PCR likewise (Wibbelt et al. 2010).
13.3.4.3 Swabs
Cotton-, polyester- or nylon-flocked swabs are equally used for sampling fungal growth from hibernating bats for culture isolation and PCR. Although immediate visual examination is not possible, swabs have been employed, for example, for quantifying the fungal load on wing membranes (Janicki et al. 2015; Langwig et al. 2015a, b; Zukal et al. 2016) via quantitative PCR (Shuey et al. 2014).
13.4