Adenohypophyseal Activity Is Controlled by Hypothalamic Releasing Hormones, Which Are Released into the Portal SystemzWhich in Turn Connects the Median Eminence of the Hypothalamus and the Anterior Pituitary Gland
Whereas neurons that compose the neurohypophysis are influenced directly by neural input within the hypothalamus, the imposition of a vascular system between the hypothalamus and the adenohypophysis requires a different type of control system.
The hypothalamus produces regulatory or hypophysiotropic hormones, which are transported to and released within the median eminence (comparable to posterior lobe hormones) (Figure 33-17). These regulatory hormones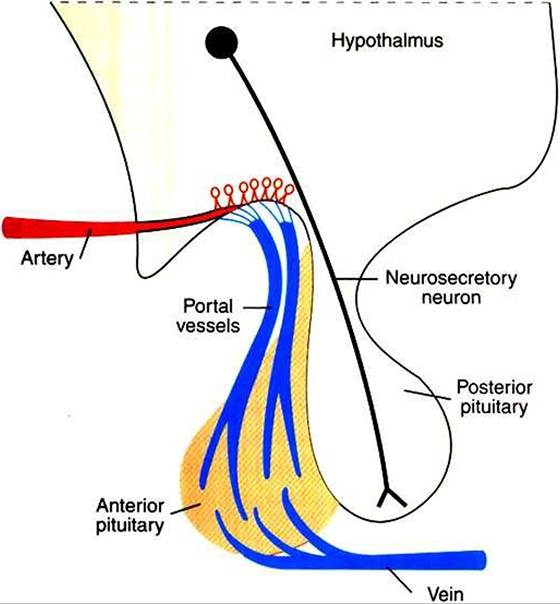
FIGURE 33-16 Diagram of the Kypothalamopituitary unit, contrasting the vascular connection between the brain and the anterior pituitary gland with the neuronal connection between the brain and the posterior pituitary gland. (From Hedge GA, Colby HD, Goodman RL: Clinical endocrine physiology, Philadelphia, 1987, Saunders.)
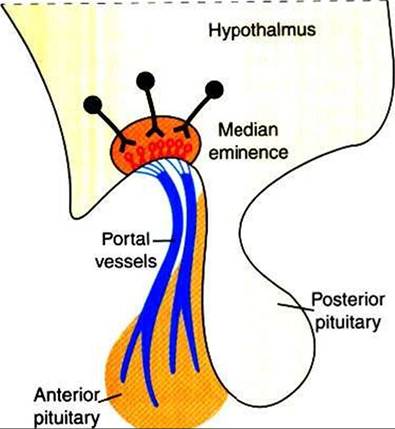
FIGURE 33-17 ■ Hypothalamic neurosecretory neurons and Hypothalamohypophyseal portal vessels. (From Hedge GA, Colby HD, Goodman RL: Clinical endocrine physiology, Philadelphia, 1987, Saunders.)
then pass via the portal venous system to the adenohypophysis, where they stimulate the release of the various anterior pituitary hormones. The synthesis of adenohypophyseal regulatory hormones is controlled by both neural and hormonal inputs at the level of the hypothalamus. Some of the hypophyseal hormones have been found in other areas of the brain and extraneural sites, including the gastrointestinal tract and the pancreas.
The initial isolation and identification of the hypothalamic hormones required large amounts of tissue as well as expertise in biochemistry. The first identified hypothalamic hormone, which controls corticotropin release, was originally called corticotropin-releasing factor (now changed from factor to hormone).
The initial work, done by Guillemins group at the University of Houston in the early 1960s, required the collection, freezing, and transport of several hundred thousand sheep brains from abattoirs located in the western United States, as well as the subsequent dissection of the hypothalami. The hypothalamic hormones that have been characterized and the hormones they release include the following (Table 33-2):1. Corticotropin-releasing hormone (CRH), a 41-amino acid polypeptide that stimulates Corticotropes to release all components of the pro-opiomelanocortin family of molecules.
2. Gonadotropin releasing hormone (GnRH), a decapeptide that stimulates gonadotrope secretion of both FSH and LH.
3. Thyrotropin-releasing hormone (TRH), a tripeptide that stimulates thyrotrope secretion of TSH.
4. Dopamine, a catecholamine precursor of norepinephrine that inhibits Iactotrope secretion of PRL and thyrotrope secretion of TSH.
5. Somatostatin, a tetradecapeptide that inhibits Somatotrope secretion of GH.
6. Growth hormone-releasing hormone (GHRH), a 44-amino acid polypeptide that stimulates Somatotrope secretion of GH.
Except for dopamine, all these hypophysiotropic hormones are peptides.
Previously, only four of the anterior pituitary hormones (FSH, LH, TSH, and corticotropin) were considered to be “tropic”; that is, their main effect was stimulation of hormone secretion by specific endocrine organs located peripheral to the pituitary gland. More recently, GH has been added to this list because GH stimulates the liver to produce somatomedins, which have a negative-feedback effect on GH secretion. PRL remains as the only pars distalis hormone for which negativefeedback inhibition has not been demonstrated through hormones produced by PRL target tissues.
The most important regulation of secretion of the protein hormones by the pars distalis is by feedback inhibition. One feedback system involves negative-feedback inhibition of the tropic pituitary hormone by interaction of the target organ hormone with the hypothalamus, as well as with
Table 33-2
Major Hypophysiotrophic Hormones
| Hormone | Abbreviation | Site of origin |
| Thyrotropin-releasing hormone | TRH | Paraventricular nucleus |
| Gonadotropin-releasing hormone | GnRH | Preoptic area of hypothalamus |
| Growth hormone-inhibiting hormone (somatostatin) | GHIH | Anterior hypothalamic area |
| Growth hormone-releasing hormone | GHRH | Arcuate nucleus |
| Corticotropin-releasing hormone | CRH | Paraventricular nucleus |
| Prolactin-releasing factor | PRF | ? |
| Prolactin-inhibiting hormone (or dopamine) | PIH | Arcuate nucleus |
Modified from Hedge GA, Colby HD, Goodman RL: Clinical endocrine physiology, Philadelphia, 1987 Saunders.
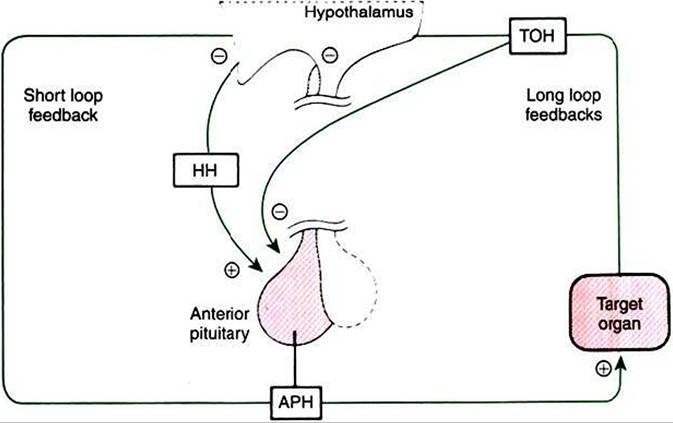
FIGURE 33-18 Regulation of anterior pituitary hormone (APH) secretion by hypophysiotropic hormones (HH)1 shortloop negative feedback, and Iong-Ioop negative feedback by target organ hormones (TOH).
Plus signs indicate stimulation, and minus signs indicate inhibition. (From Hedge GA, Colby HD, Goodman RL: Clinical endocrine physiology, Philadelphia, 1987, Saunders.)the pituitary gland; this system is called a long-loop feedback system (Figure 33-18). For example, cortisol is produced by the adrenal cortex as a result of corticotropin stimulation, and cortisol in turn has a negative-feedback effect on corticotropin production at the level of the hypothalamus and the anterior pituitary gland. Short-Ioop feedback systems have also been described; an anterior pituitary hormone such as corticotropin has a direct negative-feedback inhibition of hormone secretion, in this case CRH, within the hypothalamus.
Even under conditions of negative-feedback inhibition, the secretion of anterior pituitary hormones is not constant. For example, even though estrogens exert a continuous, potent negative-feedback inhibition on gonadotropin secretion, gonadotropin secretion alternates between secretion and no secretion, with pulses of gonadotropins released into the vascular system. In the case of gonadotropins, the ovarian endocrine status influences the pulse rate and the amplitude of each pulse. Progesterone domination is associated with a decreased pulse rate and an increased pulse amplitude, whereas estrogen causes the opposite effect. The work of Irvine and Alexander has provided the best documentation of the precise relationship between secretory activity of hypothalamic regulatory and anterior pituitary hormones. Their data were obtained through analysis of hormones obtained from the intercavernal sinus, which collects venous blood from the pituitary gland of the horse.
Clinical syndromes of somatotropin deficiency and excess include pituitary dwarfism in the dog and acromegaly in the cat, respectively. Pituitary dwarfism results from destruction of the pituitary gland through a neoplastic, degenerative, or anomalous process. It may be associated with decreased production of other pituitary hormones, including TSH, ACTH, LH, FSH, and GH.
Pituitary dwarfism is most common in German shepherd dogs aged 2 to 6 months. Other affected breeds include Carnelian bear dogs, spitz, toy pinschers, and Weimaraners. In German shepherd dogs the disease is inherited as a simple autosomal recessive trait and occurs as a result of cystic Rathke’s pouch. The first observable clinical signs of pituitary dwarfism are slow growth, noticed in the first 2 to 3 months of life, and mental retardation, usually manifested as difficulty in house training. Physical examination findings may include proportionate dwarfism, retained puppy hair coat, hypotonic skin, truncal alopecia, cutaneous hyperpigmentation, infantile genitalia, and delayed dental eruption. Clinicopathological features include eosinophilia, lymphocytosis, mild normocytic-normochromic anemia, hypophosphatemia, and occasionally hypoglycemia resulting from secondary adrenal insufficiency. Differential diagnoses include other causes of stunted growth, such as hypothyroid dwarfism, portosystemic shunt, diabetes mellitus, hyper- adrenocorticism, malnutrition, and parasitism. Diagnosis is made by measuring serum growth hormone concentrations (assay no longer commercially available) or serum somatomedin C (insulin-like growth factor 1 [IGF-1∣). The advantage of IGF-I is that it is not species specific. There is usually a subnormal response to exogenous TSH and ACTH stimulation tests; furthermore, endogenous TSH and ACTH are decreased in affected dogs as a result of panhypopituitarism.Acromegalyt or Iiypersomatotropismt is the condition resulting from chronic excessive GH secretion in the adult animal. Canine acromegaly is an extremely rare disorder observed after the administration of progestational compounds for suppression of estrus in intact female dogs. The disease is caused by excessive secretion of GH from mammary cells under the influence of exogenous progesterone. Acromegaly in cats, as in humans, is caused by a GH-secreting tumor of the anterior pituitary gland.
Such tumors in cats grow slowly and may be present for a long time before the onset of clinical signs. Feline acromegaly occurs in older (8- to 14-year-old) cats and occurs more frequently in males. Canine acromegaly occurs in intact female dogs given progestational compounds for estrus prevention.Clinical signs of uncontrolled diabetes mellitus (DM) are often observed as the first manifestations of acromegaly; therefore polydipsia, polyuria, and polyphagia are the most common presenting signs. Net weight gain of lean body mass in animals with uncontrolled DM is a key sign of acromegaly. Organomegaly, including renomegaly (observed in both cats and humans with acromegaly), hepatomegaly, and enlargement of endocrine organs, is also observed. Some dogs and cats show the classic enlargement of extremities, body size, jaw, tongue, and forehead that is characteristic of acromegaly in humans. Some of the most striking manifestations of acromegaly occur in the musculoskeletal system, such as an increase in muscle mass and growth of the acral segments of the body, including the paws, chin, and skull. Cardiovascular abnormalities, such as Cardiomegaly (as determined radiographically and echocardiographically), systolic murmurs, and congestive heart failure, develop late in the course of the disease. Azotemia develops late in the disease in approximately 50% of acromegalic cats. Neurological signs of acromegaly in humans, such as peripheral neuropathies (paresthesias, carpal tunnel syndrome, sensory and motor defects), and parasellar manifestations, such as headache and visual field defects, are not generally detected in acromegalic small animals.
Impairments in glucose tolerance and insulin resistance that result in DM are observed in all cats and most dogs with acromegaly. Measurement of endogenous insulin reveals dramatically increased serum insulin concentrations. Despite severe insulin resistance and hyperglycemia, ketosis is rare in acromegalic animals. Feline acromegaly should be suspected in any diabetic cat (especially males) that has severe insulin resistance (insulin requirement >20 L∣∕cat∕day).
Hypercholesterolemia and mild increases in serum activities of liver enzymes are attributed to the diabetic state. Hyperphosphatemia without azotemia is also a common Clinicopathological finding, perhaps as a result of GH-StimuIated bone growth. Urinalysis findings are unremarkable except for persistent proteinuria, probably as a result of systemic hypertension and glomerulosclerosis.A definitive diagnosis of acromegaly requires documentation of increased plasma G or somatomedin C concentrations. Unfortunately, feline and canine GH assays are no longer commercially available.
Cushings syndrome and hyperthyroidism should be ruled out before measurement of GH. At this time the most definitive test for the diagnosis of acromegaly in cats is computed tomography (CT) of the pituitary region. CT findings, coupled with the exclusion of other disorders that cause insulin resistance (hyperthyroidism, Iiyperadrenocorticism), in cats that exhibit clinical signs of acromegaly should lead the clinician to a diagnosis of acromegaly.