Plasma Osmolality Controls the Secretion of Vasopressin
The control of vasopressin secretion as a result of changes in plasma osmolality is through osmoreceptors located in the hypothalamus as well as through receptors located in the esophagus and stomach that immediately sense water intake (Figure 33-14).
An increase in osmolality of body fluids increases the rate of action potential firing in the osmoreceptors, which in turn activates hypothalamic cells that synthesize vasopressin. This negative-feedback system is sensitive to changes in osmolality, and the solute-to-water ratio is maintained within 1% to 2% of the normal values. The regulation of the pressor effect of vasopressin—that is, through blood volume— is achieved by increasing the number of action potentials in stretch receptors located in the atria. A decrease in blood volume activates the stretch receptors, which inhibit activity of neurons, vagal in origin, that inhibit the osmoreceptor cells. Blood volume changes that decrease blood pressure also affect vasopressin release through activation of baroreceptors in the carotid sinus and aortic arch.Diabetes insipidus (DI) is a disorder of water metabolism characterized by polyuria, urine of low specific gravity or osmolality, and polydipsia. It is caused by defective secretion of ADH (central DI) or by the inability of the renal tubule to respond to ADH (nephrogenic DI). Deficiency of ADH can be partial or complete. Central DI is characterized by an absolute or relative lack of circulating ADH and is classified as primary (idiopathic and congenital) or secondary. Secondary central DI usually results from head trauma or neoplasia.
Central DI may appear at any age, in any breed of dog or cat, and in either gender; however, young adults (6 months of age) are most often affected. The major clinical signs of DI are profound polyuria and polydipsia OlOO mL/kg/day; normal range, 40-70 mL/kg/day), nocturia, and incontinence, usually of several months’ duration.
The severity of the clinical signs varies because DI may result from a partial or complete defect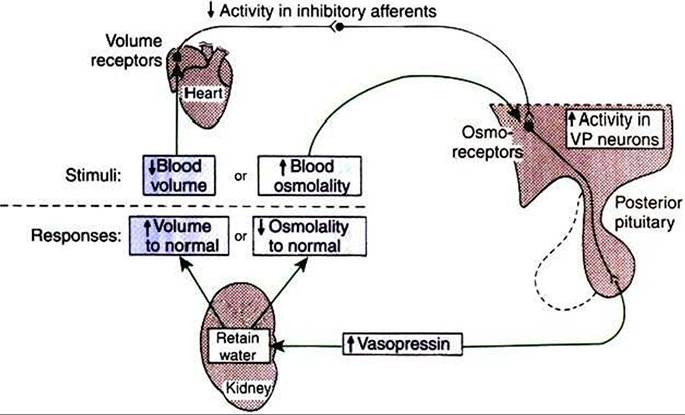
FIGURE 33-14 Major mechanisms regulating vasopressin (VP) secretion. A perturbation in either blood volume or osmolality modifies vasopressin secretion to restore these parameters to their normal values. However, this restoration requires appropriate water intake adjustments by thirst as well as by the modulation of water retention depicted. Also, the two responses indicated may be affected by simultaneous changes in sodium balance. (From Hedge GA, Colby HD, Goodman RL: Clinical endocrine physiology, Philadelphia, 1987, Saunders.)
in ADH secretion or action. Other, less consistent signs include weight loss (because these animals are constantly seeking water) and dehydration.
Routine complete blood cell count and serum biochemical and electrolyte profiles are usually normal in animals with DI. Plasma osmolality is often high (>310 mOsm∕L) in central or nephrogenic Dl as a result of dehydration. Animals with primary polydipsia often exhibit low plasma osmolality (mare’s serum gonadotropin”) and primate chorionic gonadotropin, which are produced by cells of the placental chorion.
Corticotropin and β-lipotropin belong to the proopiomelanocortin family in that they originate from a common prohormone (Figure 33-15). Cells in both the pars distalis and the pars intermedia synthesize proopiomelanocortin molecules. The emphasis on the type of hormone produced is different in the end product; corticotropin is produced by pars distalis Corticotropes. In the pars intermedia, corticotropin is cleaved by Corticotropes to form Ct-Welanocyte-Stimulating hormone (α-MSH), the predominant hormone of this lobe. The remaining peptide fragment is known as corticotropin-like intermediate lobe peptide; the physiological activity of this peptide fragment is not known.
In both the pars distalis and the pars intermedia, β-lipotropin is cleaved to form β-endorphins and γ-lipotropin. Endorphins have opioid activity and appear to modulate gonadotropin secretion.Control of adenohypophyseal activity was not understood for many years, primarily because the functional connection between the brain and the anterior pituitary gland was not understood. In the 1930s, Popa and Fielding, Budapest medical student and university professor, respectively, described the vascular system that connects the hypothalamus with the pituitary gland but were unable to determine the direction in which blood flowed. In about 1950, Geoffrey Harris drew the important conclusion that the linkage involved blood passage from the hypothalamus to the anterior pituitary gland through the portal blood system previously described by Popa and Fielding (Figure 33-16). The dorsal hypophyseal artery, which supplies nutrients and oxygen to the adenohypophysis (the ventral hypophyseal artery supplies the neurohypophysis), terminates in the median eminence as a capillary plexus. Blood from these plexus is drained by two veins that empty into sinusoidal capillaries of the pars distalis, completing the portal venous system (one vein supplies the ventral, central part of the pars distalis; the other supplies the dorsal, peripheral areas).