Age and Functional Changes
Although there has been little detailed study of the postnatal development, it is known that the testes most often remain within the abdomen until about the third day after birth.
Their descent through the inguinal canal then commences, and although it is completed within a couple of days, another 4 or 5 weeks is required before the testes occupy their definitive positions within the scrotum. The seminiferous tissue increases markedly in volume during this time, but spermatogenesis does not begin until about the sixth month. Because the testes attain their definitive locations so precociously, some have advocated castration of male kittens at much younger ages—6 to 14 weeks—rather than the 5 or 6 months conventionally adopted. It is claimed that the operation is well tolerated by these very young animals.* If descent fails—the cryptorchid condition—the testis may be located anywhere between the caudal pole of the kidney and the inguinal canal. It is most easily located by following the deferent duct, which is readily picked up at the lateral ligament of the bladder. Although the germinal epithelium fails to develop normally at the core temperature of the body, Leydig cells produce androgens, and the full range of secondary sex characteristics may develop in bilaterally cryptorchid animals.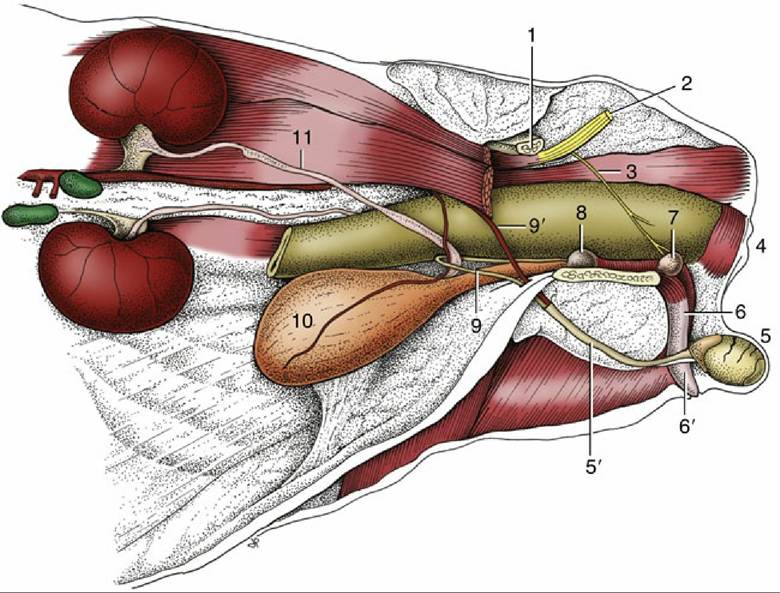
FIG. 15.22 The reproductive organs of the tomcat in situ, left lateral view. 1, Shaft of ilium; 2, sciatic nerve; 3, pudendal nerve; 4, anus; 5, left testis in scrotum; 5', spermatic cord; 6, penis; 6', prepuce; 7, bulbourethral gland; 8, prostate; 9, deferent duct; 9', testicular vessels; 10, bladder; 11, left ureter.
The mating behavior of dogs is most unusual. The dog mounts the bitch in the usual way, but shortly after intromission he drops to her side and reverses so that the pair stand rear to rear during the remainder of the "tie," which may last for a further 45 minutes or even longer.
There has been surprisingly little consideration of the anatomy of this process.Although all erectile tissues of the penis become engorged when erection is complete, they attain very different degrees of expansion and turgidity (Fig. 15.20). The corpus cavernosum swells least, and its construction allows it to remain flexible about a vertical axis, though not about a horizontal one, even in this state. The bulbus glandis is most capable of expansion and swells to twice its resting thickness, becoming very tense in the process. The pars longa stiffens least but elongates considerably, causing it to slide apically on the os penis to which it is only loosely attached. It then extends well beyond the fibrocartilaginous extension of the bone and presents an indentation about the urethral orifice in consequence of the tighter anchorage of this part.
Intromission necessarily occurs before the penis is markedly enlarged (Fig. 15.21D and E). The labia thrust the prepuce caudally when the dog mounts and introduces the glans into the vagina. The slope of the female passage requires a dorsocranial penetration, and the relatively soft tip of the glans is diverted ventrally by its impingement (through the soft tissues) on the pelvic roof. This deflection allows the penis to be advanced toward the fornix and perhaps explains the necessity for the softer nature of the pars longa and the early termination of its bony support. When the stud dog dismounts and turns through 180 degrees, the body of the penis is bent laterally and then caudally; withdrawal of the penis is prevented by the swollen bulbus glandis and the grip exerted on it by the engorged vestibular bulbs and muscles associated with the female tract. The reversal of position twists the prepuce, tightening the preputial muscles into a cord that presses on the veins draining the glans. The dorsal veins of the penis, buckled by the flexion of the penis, are further obstructed by being pressed against the ischial arch by the contraction of the ischiourethralis.
Detumescence is probably eventually achieved by relaxation of the bulbospongiosus, which allows the spaces within the corpus spongiosum to provide alternative channels for the escape of blood from the engorgedpenis.
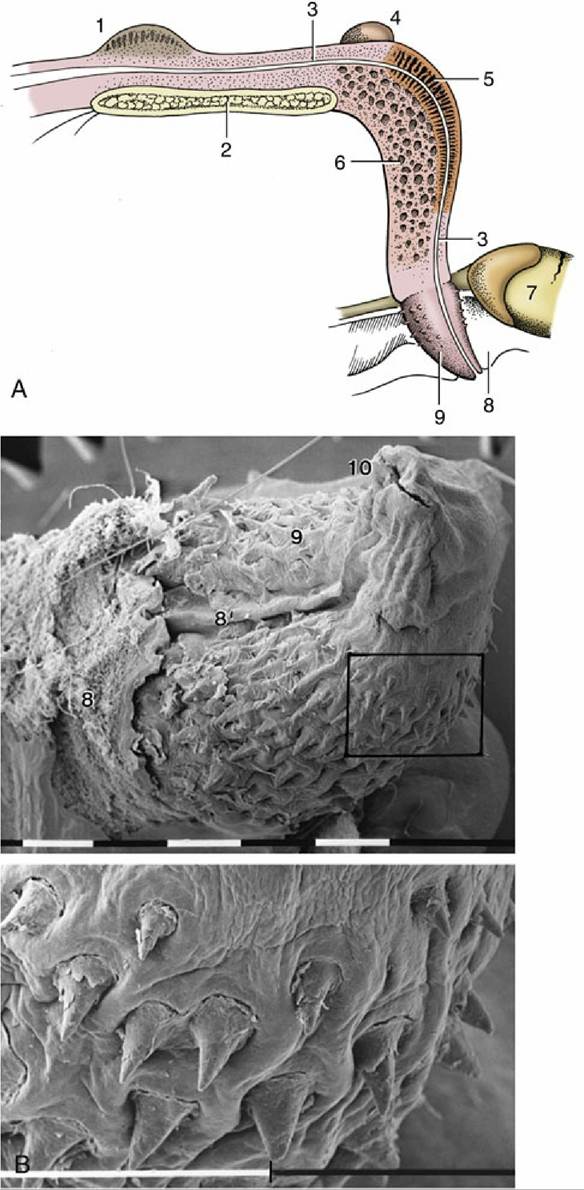
FIG. 15.23 (A) Median section of the feline penis, left lateral view. (B) Scanning electron micrograph of a feline glans and enlargement of the marked area (bar = 1 mm). 1, Prostate; 2, pelvic symphysis; 3, urethra; 4, right bulbourethral gland; 5, corpus spongiosum; 6, corpus cavernosum; 7, right testis; 8, prepuce; 8', preputial frenulum; 9, glans (with spines); 10, external urethral orifice.
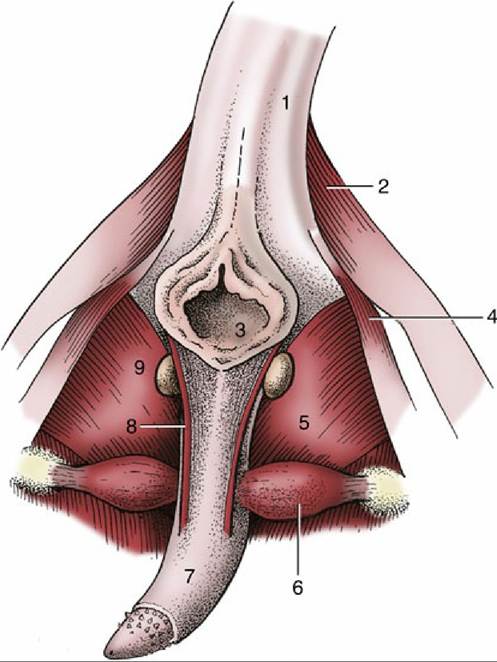
FIG. 15.24 Feline penis in situ, caudal view. 1, Tail (raised); 2, gluteofemoralis muscle; 3, anus; 4, coccygeus muscle; 5, internal obturator muscle; 6, ischiocavernosus muscle; 7, penis; 8, left retractor penis; 9, left bulbourethral gland.

FIG. 15.25 Tomcat spraying.
The initial, sperm-rich fraction of the ejaculate is discharged during the first stage of coitus when the dog is mounted in the fashion conventional for quadrupeds. The second stage is occupied in pumping out the much larger fraction—perhaps 30 mL—provided by the prostate; the tide sweeps the sperm-rich part through the cervix into the body of the uterus. It is known that short matings — in which only first-stage coitus occurs—may be fertile. The purpose of the second-stage coitus may encourage uterine rather than vaginal insemination. Turning around discourages detumescence of the penis and therefore maintains high intravaginal pressure.

FIG. 15.26 Mating posture.
The penis of the cat increases considerably in length on erection and then curves downward and forward. This change in orientation, allied to a ventral flexion of the pelvic region, enables coitus to be performed in a fashion not greatly different from that usual in quadrupeds (Fig. 15.26).